

GNA13 loss in germinal center B cells leads to impaired apoptosis and promotes lymphoma in vivo
- PMID: 26989201
- PMCID: PMC4991089
- DOI: 10.1182/blood-2015-07-659938
GNA13 loss in germinal center B cells leads to impaired apoptosis and promotes lymphoma in vivo
Abstract
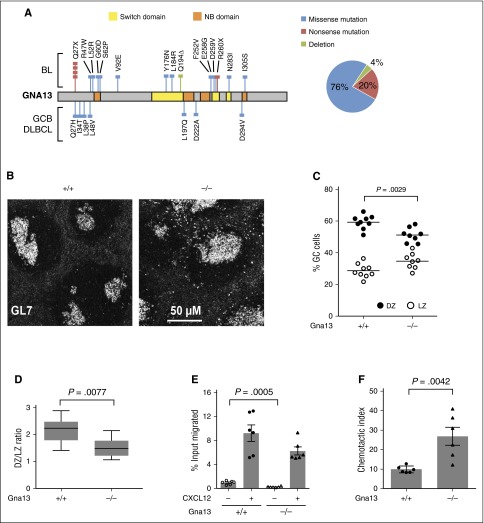
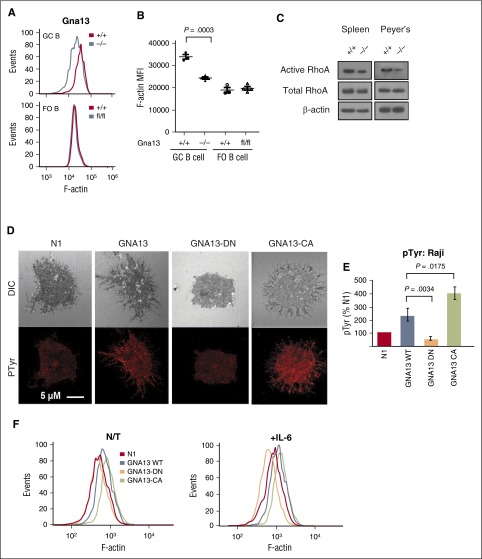
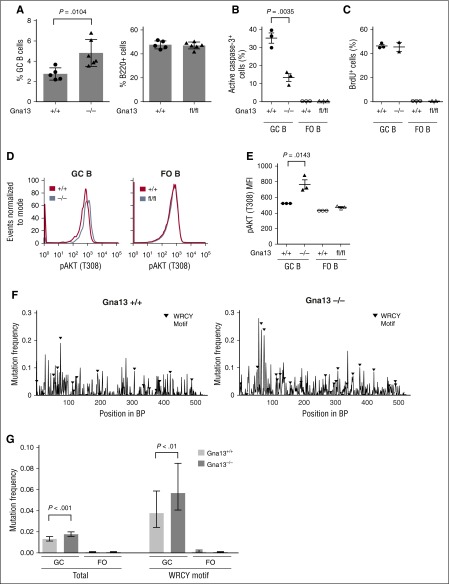
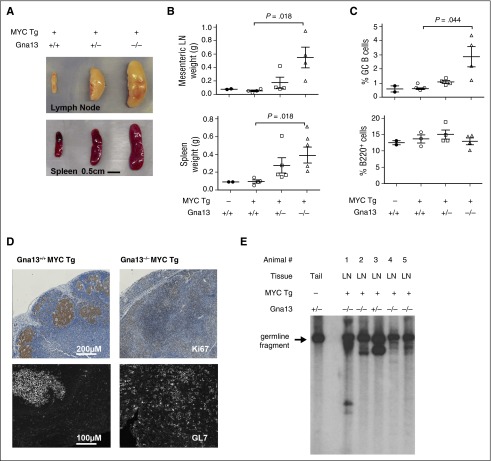
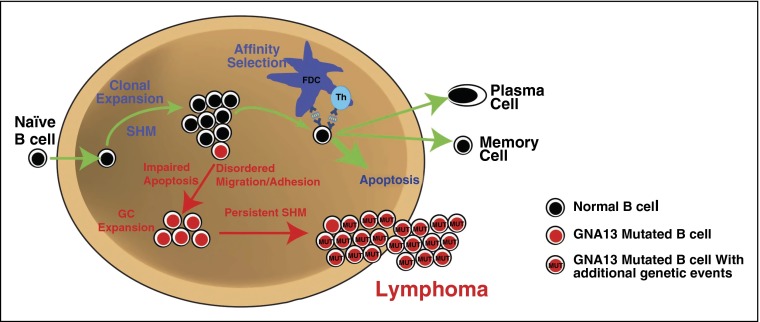
GNA13 is the most frequently mutated gene in germinal center (GC)-derived B-cell lymphomas, including nearly a quarter of Burkitt lymphoma and GC-derived diffuse large B-cell lymphoma. These mutations occur in a pattern consistent with loss of function. We have modeled the GNA13-deficient state exclusively in GC B cells by crossing the Gna13 conditional knockout mouse strain with the GC-specific AID-Cre transgenic strain. AID-Cre(+) GNA13-deficient mice demonstrate disordered GC architecture and dark zone/light zone distribution in vivo, and demonstrate altered migration behavior, decreased levels of filamentous actin, and attenuated RhoA activity in vitro. We also found that GNA13-deficient mice have increased numbers of GC B cells that display impaired caspase-mediated cell death and increased frequency of somatic hypermutation in the immunoglobulin VH locus. Lastly, GNA13 deficiency, combined with conditional MYC transgene expression in mouse GC B cells, promotes lymphomagenesis. Thus, GNA13 loss is associated with GC B-cell persistence, in which impaired apoptosis and ongoing somatic hypermutation may lead to an increased risk of lymphoma development.
© 2016 by The American Society of Hematology.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
