

C9orf72 is required for proper macrophage and microglial function in mice
- PMID: 26989253
- PMCID: PMC5120541
- DOI: 10.1126/science.aaf1064
C9orf72 is required for proper macrophage and microglial function in mice
Abstract
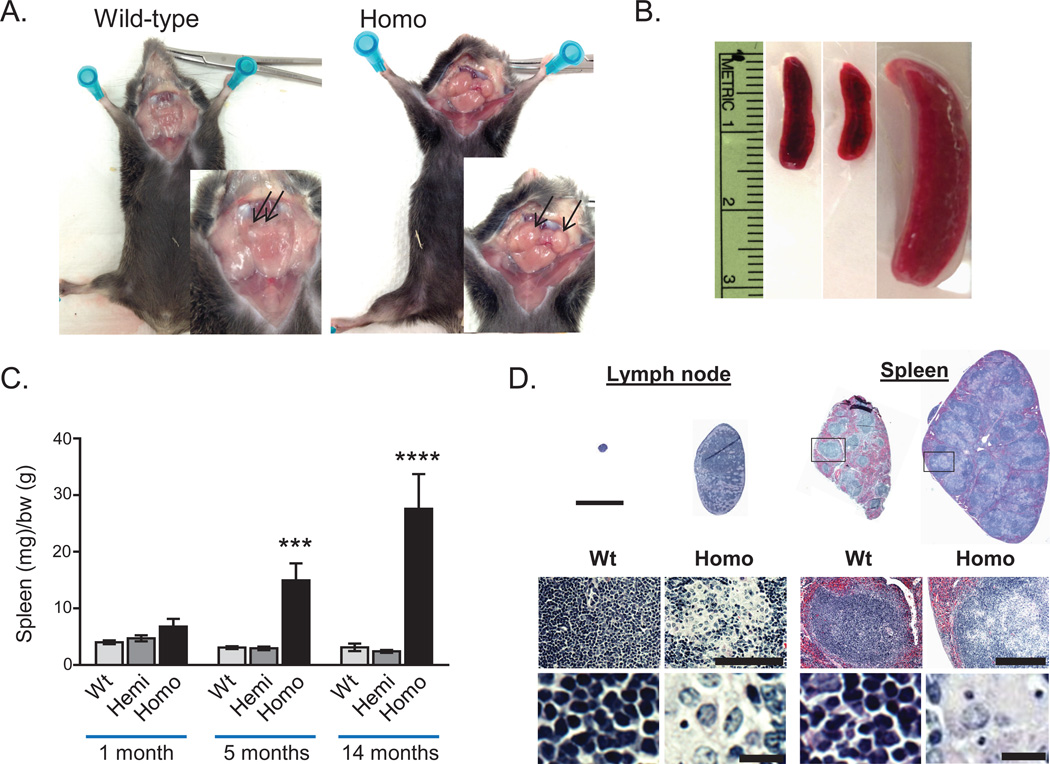
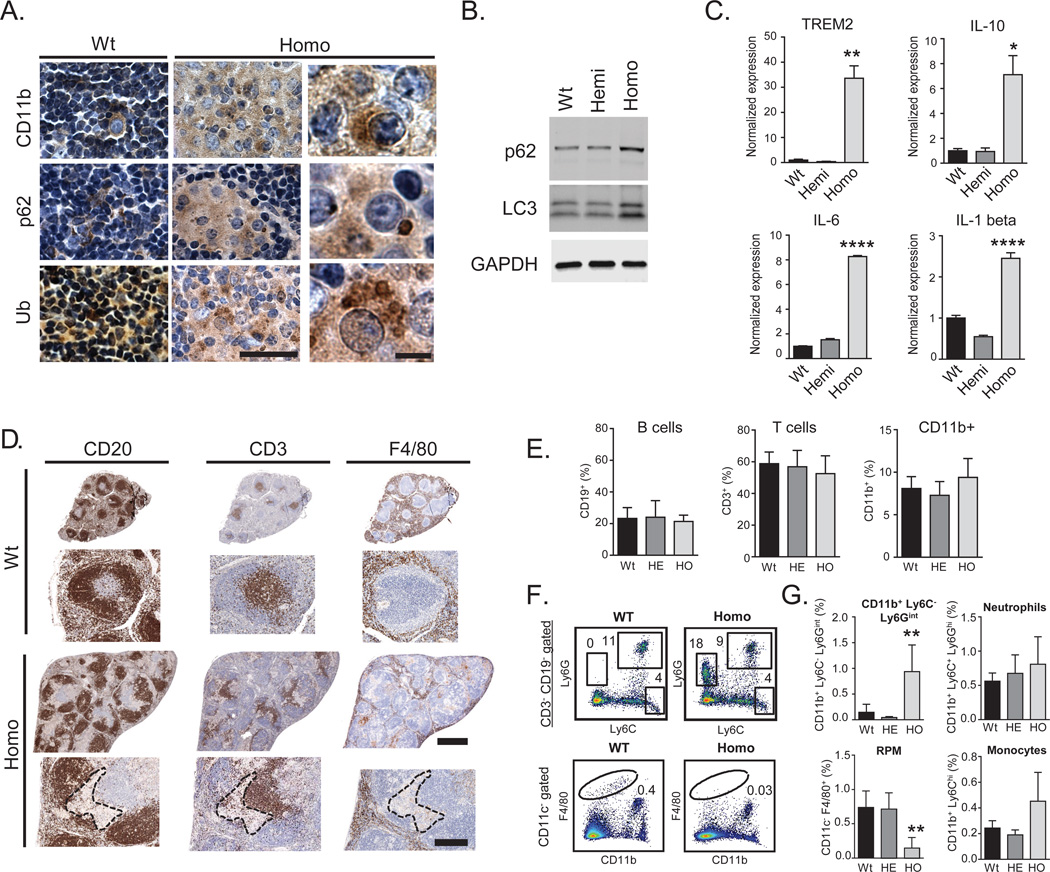
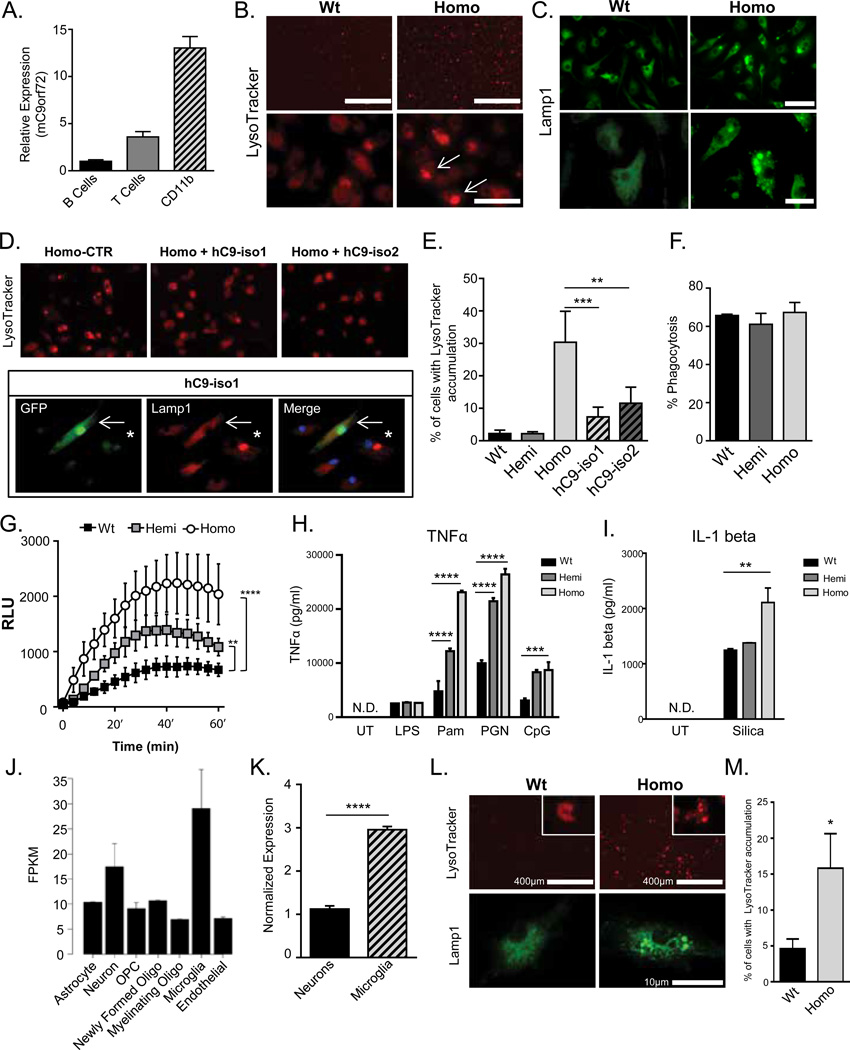
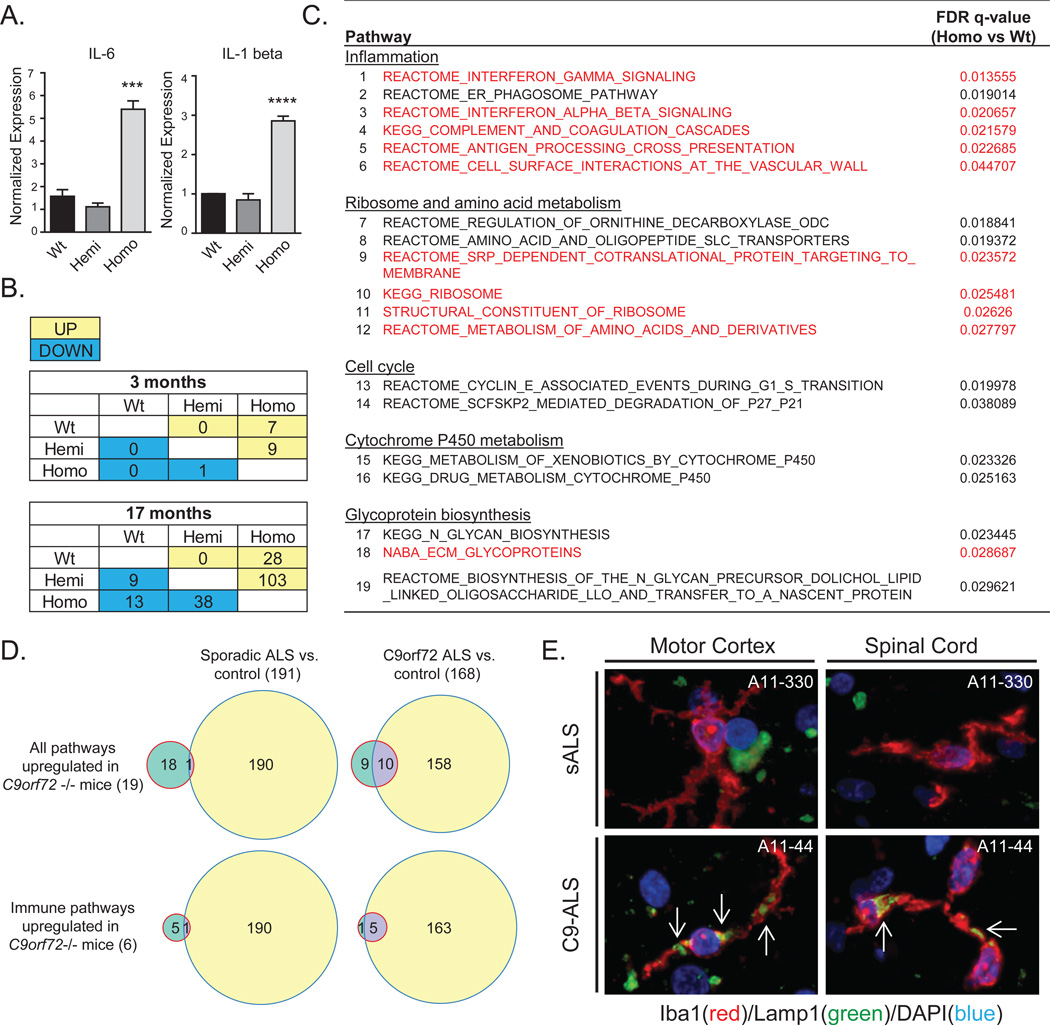
Expansions of a hexanucleotide repeat (GGGGCC) in the noncoding region of the C9orf72 gene are the most common genetic cause of amyotrophic lateral sclerosis (ALS) and frontotemporal dementia. Decreased expression of C9orf72 is seen in expansion carriers, suggesting that loss of function may play a role in disease. We found that two independent mouse lines lacking the C9orf72 ortholog (3110043O21Rik) in all tissues developed normally and aged without motor neuron disease. Instead, C9orf72 null mice developed progressive splenomegaly and lymphadenopathy with accumulation of engorged macrophage-like cells. C9orf72 expression was highest in myeloid cells, and the loss of C9orf72 led to lysosomal accumulation and altered immune responses in macrophages and microglia, with age-related neuroinflammation similar to C9orf72 ALS but not sporadic ALS human patient tissue. Thus, C9orf72 is required for the normal function of myeloid cells, and altered microglial function may contribute to neurodegeneration in C9orf72 expansion carriers.
Copyright © 2016, American Association for the Advancement of Science.
Figures
Comment in
-
Full ablation of C9orf72 in mice causes immune system-related pathology and neoplastic events but no motor neuron defects.Acta Neuropathol. 2016 Jul;132(1):145-7. doi: 10.1007/s00401-016-1581-x. Epub 2016 May 20. Acta Neuropathol. 2016. PMID: 27206760 Free PMC article. No abstract available.
References
-
- Lomen-Hoerth C, Anderson T, Miller B. The overlap of amyotrophic lateral sclerosis and frontotemporal dementia. Neurology. 2002 Oct 8;59:1077. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
