

Transient Surface CCR5 Expression by Naive CD8+ T Cells within Inflamed Lymph Nodes Is Dependent on High Endothelial Venule Interaction and Augments Th Cell-Dependent Memory Response
- PMID: 26994221
- PMCID: PMC4868656
- DOI: 10.4049/jimmunol.1501176
Transient Surface CCR5 Expression by Naive CD8+ T Cells within Inflamed Lymph Nodes Is Dependent on High Endothelial Venule Interaction and Augments Th Cell-Dependent Memory Response
Abstract
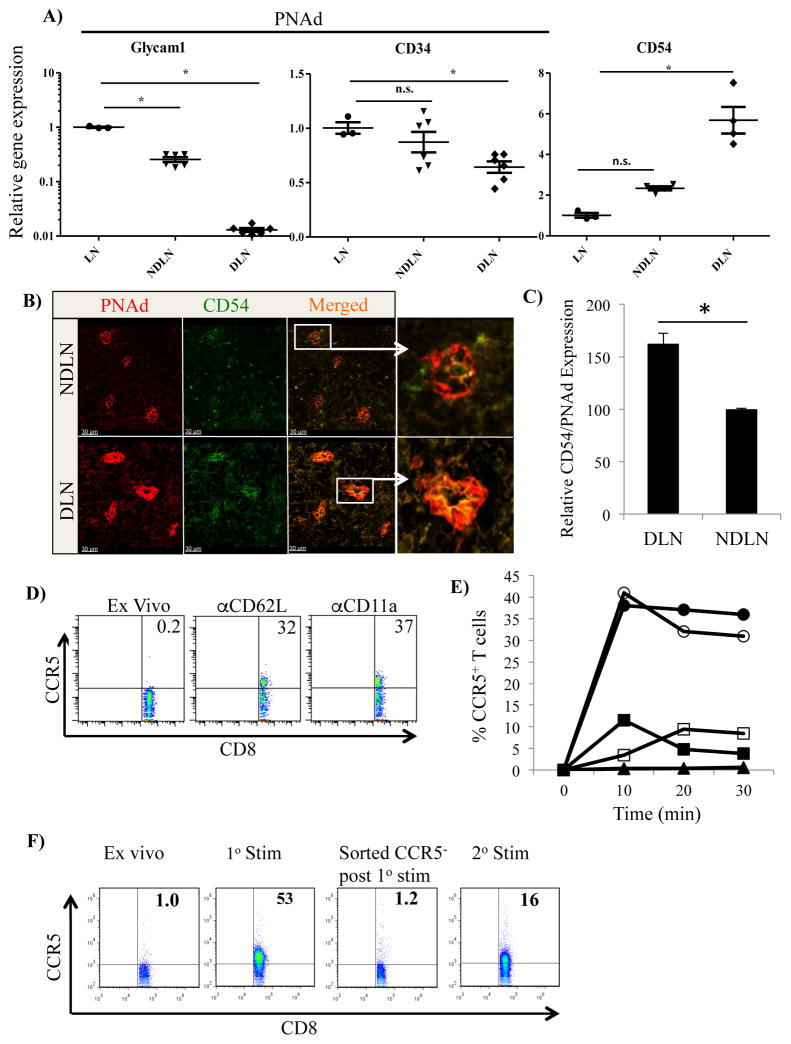
In inflamed lymph nodes, Ag-specific CD4(+) and CD8(+) T cells encounter Ag-bearing dendritic cells and, together, this complex enhances the release of CCL3 and CCL4, which facilitate additional interaction with naive CD8(+) T cells. Although blocking CCL3 and CCL4 has no effect on primary CD8(+) T cell responses, it dramatically impairs the development of memory CD8(+) T cells upon Ag rechallenge. Despite the absence of detectable surface CCR5 expression on circulating native CD8(+) T cells, these data imply that naive CD8(+) T cells are capable of expressing surface CCR5 prior to cognate Ag-induced TCR signaling in inflamed lymph nodes; however, the molecular mechanisms have not been characterized to date. In this study, we show that CCR5, the receptor for CCL3 and CCL4, can be transiently upregulated on a subset of naive CD8(+) T cells and that this upregulation is dependent on direct contact with the high endothelial venule in inflamed lymph node. Binding of CD62L and CD11a on T cells to their ligands CD34 and CD54 on the high endothelial venule can be enhanced during inflammation. This enhanced binding and subsequent signaling promote the translocation of CCR5 molecules from intracellular vesicles to the surface of the CD8(+) T cell. The upregulation of CCR5 on the surface of the CD8(+) T cells increases the number of contacts with Ag-bearing dendritic cells, which ultimately results in increased CD8(+) T cell response to Ag rechallenge.
Copyright © 2016 by The American Association of Immunologists, Inc.
Figures




References
-
- Castellino F, Germain RN. Cooperation between CD4+ and CD8+ T cells: when, where, and how. Annual review of immunology. 2006;24:519–540. - PubMed
-
- Castellino F, Huang AY, Altan-Bonnet G, Stoll S, Scheinecker C, Germain RN. Chemokines enhance immunity by guiding naive CD8+ T cells to sites of CD4+ T cell-dendritic cell interaction. Nature. 2006;440:890–895. - PubMed
-
- Hugues S, Scholer A, Boissonnas A, Nussbaum A, Combadiere C, Amigorena S, Fetler L. Dynamic imaging of chemokine-dependent CD8+ T cell help for CD8+ T cell responses. Nature immunology. 2007;8:921–930. - PubMed
-
- von Andrian UH, Mempel TR. Homing and cellular traffic in lymph nodes. Nat Rev Immunol. 2003;3:867–878. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
