

Roles for lipid heterogeneity in immunoreceptor signaling
- PMID: 26995463
- PMCID: PMC5319924
- DOI: 10.1016/j.bbalip.2016.03.019
Roles for lipid heterogeneity in immunoreceptor signaling
Abstract
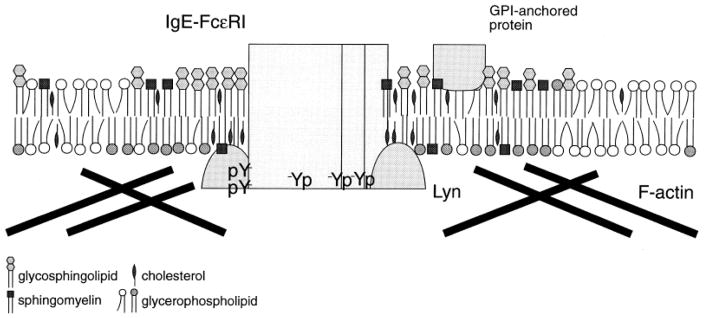
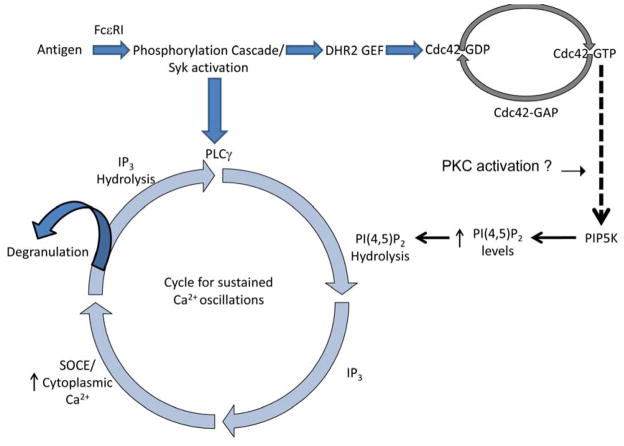
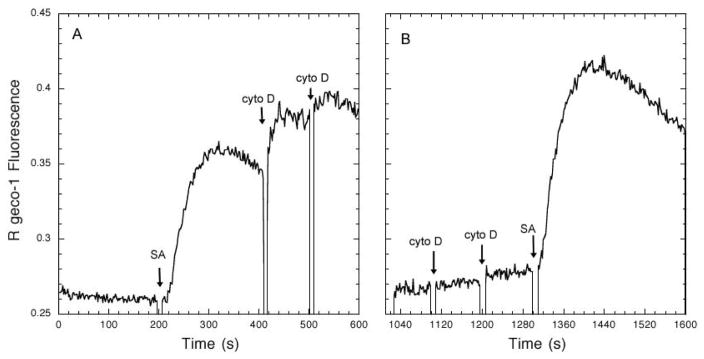
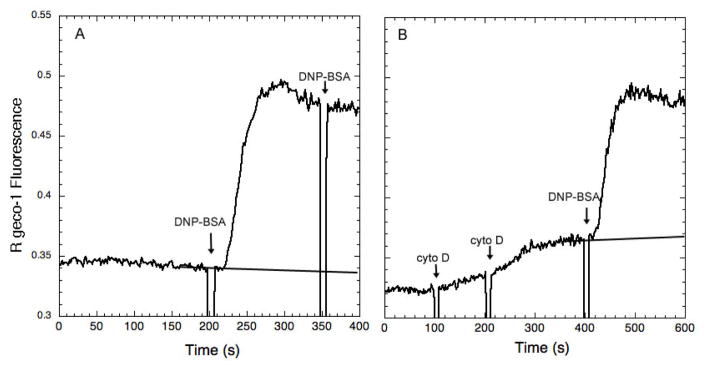
Immune receptors that specifically recognize foreign antigens to activate leukocytes in adaptive immune responses belong to a family of multichain cell surface proteins. All of these contain immunoreceptor tyrosine-based activation motifs in one or more subunits that initiate signaling cascades following stimulated tyrosine phosphorylation by Src-family kinases. As highlighted in this review, lipids participate in this initial activation step, as well as in more downstream signaling steps. We summarize evidence for cholesterol-dependent ordered lipids serving to regulate the store-operated Ca(2+) channel, Orai1, and we describe the sensitivity of Orai1 coupling to the ER Ca(2+) sensor, STIM1, to inhibition by polyunsaturated fatty acids. Phosphoinositides play key roles in regulating STIM1-Orai1 coupling, as well as in the stimulated Ca(2+) oscillations that are a consequence of IgE receptor signaling in mast cells. They also participate in the coupling between the plasma membrane and the actin cytoskeleton, which regulates immune receptor responses in T cells, B cells, and mast cells, both positively and negatively, depending on the cellular context. Recent studies show that other phospholipids with mostly saturated acylation also participate in coupling between receptors and the actin cytoskeleton. Lipid heterogeneity is a central feature of the intimate relationship between the plasma membrane and the actin cytoskeleton. The detailed nature of these interactions and how they are dynamically regulated to initiate and propagate receptor-mediated cell signaling are challenging questions for further investigation. This article is part of a Special Issue entitled: The cellular lipid landscape edited by Tim P. Levine and Anant K. Menon.
Keywords: Actin cytoskeleton; Calcium mobilization; Critical fluctuations; IgE receptors; Liquid ordered lipids; Phosphoinositides.
Copyright © 2016. Published by Elsevier B.V.
Figures
References
-
- Parham P. The Immune System. 4. Garland Science; New York, New York: 2014.
-
- van der Merwe PA, Davis SJ. Molecular interactions mediating T cell antigen recognition. Annual Review of Immunology. 2003;21:659–684. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
