

Reprogramming eukaryotic translation with ligand-responsive synthetic RNA switches
- PMID: 26999002
- PMCID: PMC4850110
- DOI: 10.1038/nmeth.3807
Reprogramming eukaryotic translation with ligand-responsive synthetic RNA switches
Abstract
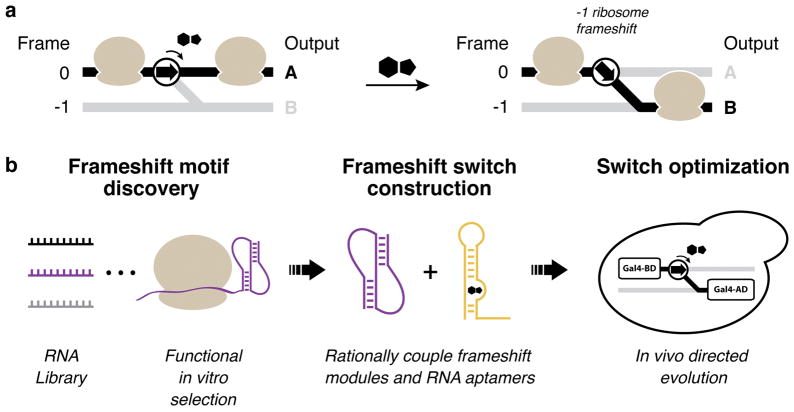
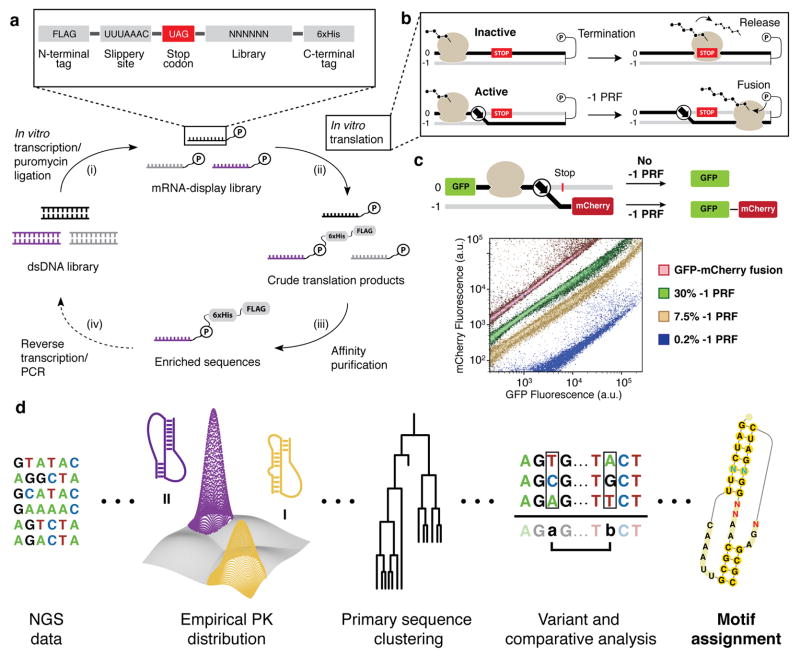
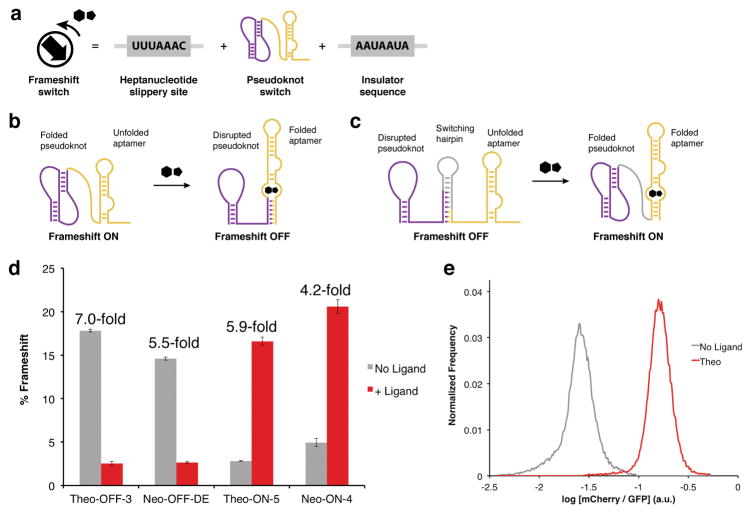
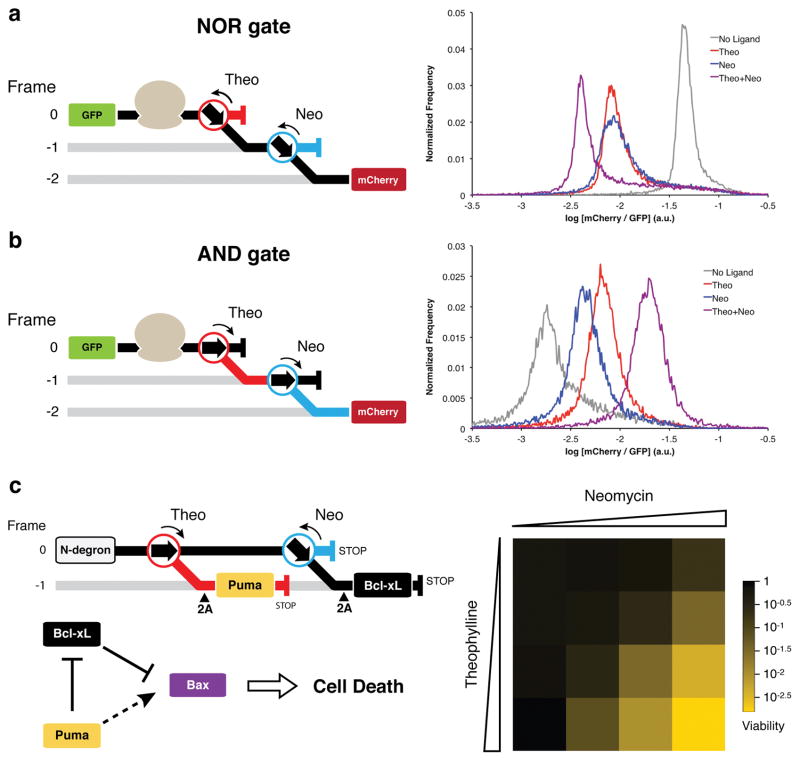
Protein synthesis in eukaryotes is regulated by diverse reprogramming mechanisms that expand the coding capacity of individual genes. Here, we exploit one such mechanism, termed -1 programmed ribosomal frameshifting (-1 PRF), to engineer ligand-responsive RNA switches that regulate protein expression. First, efficient -1 PRF stimulatory RNA elements were discovered by in vitro selection; then, ligand-responsive switches were constructed by coupling -1 PRF stimulatory elements to RNA aptamers using rational design and directed evolution in Saccharomyces cerevisiae. We demonstrate that -1 PRF switches tightly control the relative stoichiometry of two distinct protein outputs from a single mRNA, exhibiting consistent ligand response across whole populations of cells. Furthermore, -1 PRF switches were applied to build single-mRNA logic gates and an apoptosis module in yeast. Together, these results showcase the potential for harnessing translation-reprogramming mechanisms for synthetic biology, and they establish -1 PRF switches as powerful RNA tools for controlling protein synthesis in eukaryotes.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
References
-
- Gesteland RF, Atkins JF. Recoding: Dynamic Reprogramming of Translation. Annu Rev Biochem. 1996;65:741–768. - PubMed
-
- Atkins JF, Gesteland RF. Recoding: Expansion of Decoding Rules Enriches Gene Expression. Springer; New York: 2010.
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
