

Loss of the calcium channel β4 subunit impairs parallel fibre volley and Purkinje cell firing in cerebellum of adult ataxic mice
- PMID: 27003325
- PMCID: PMC4949674
- DOI: 10.1111/ejn.13241
Loss of the calcium channel β4 subunit impairs parallel fibre volley and Purkinje cell firing in cerebellum of adult ataxic mice
Abstract
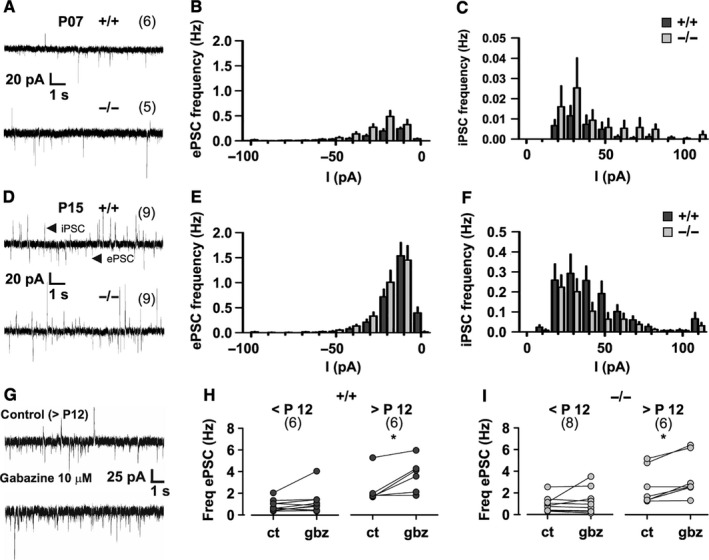
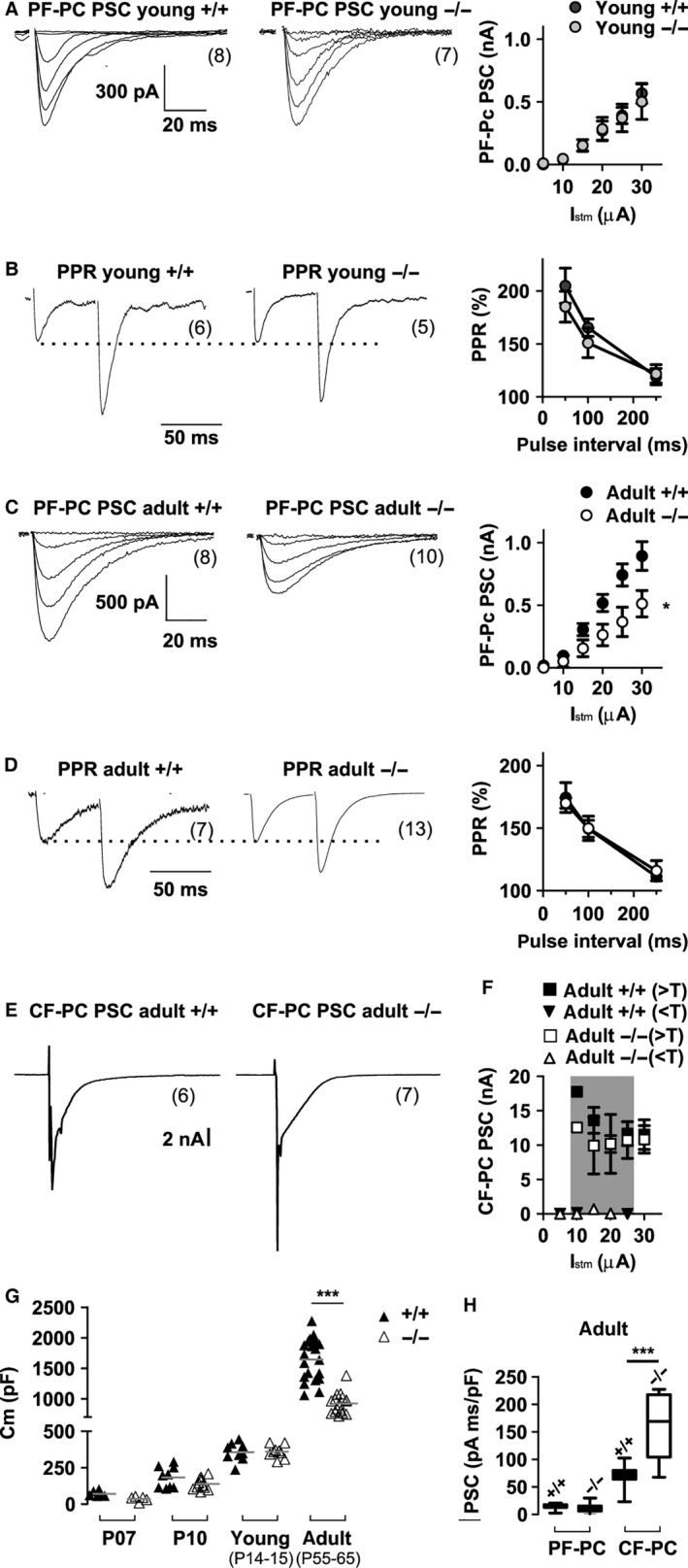
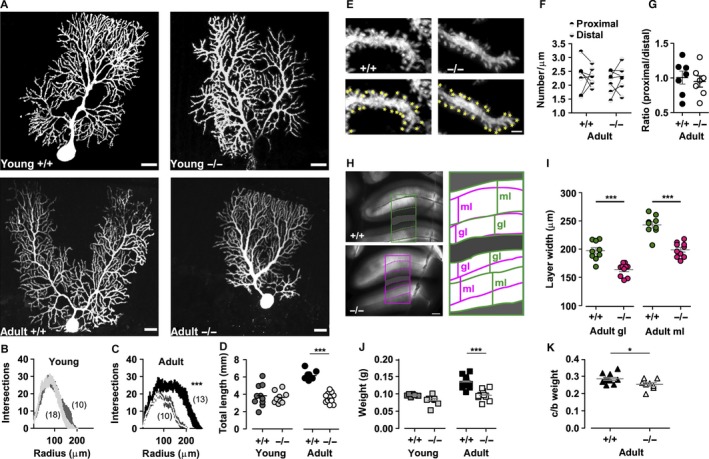
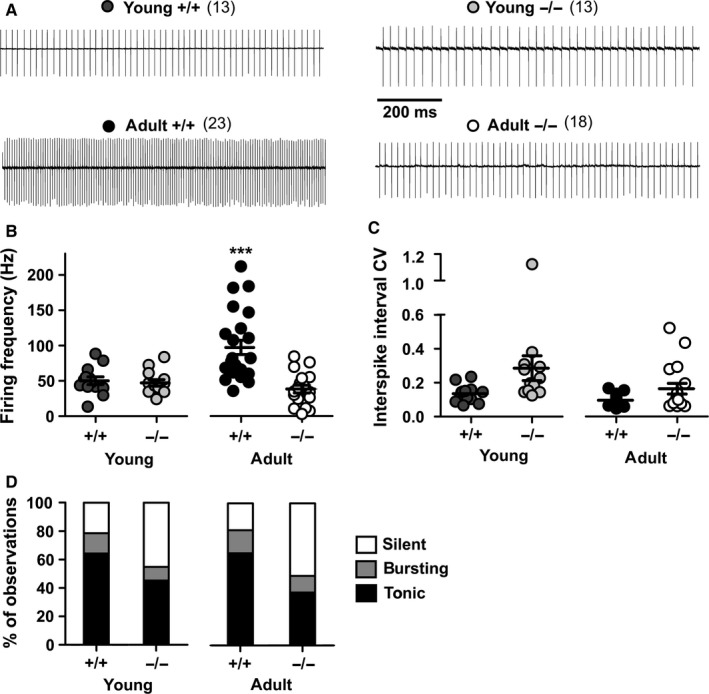
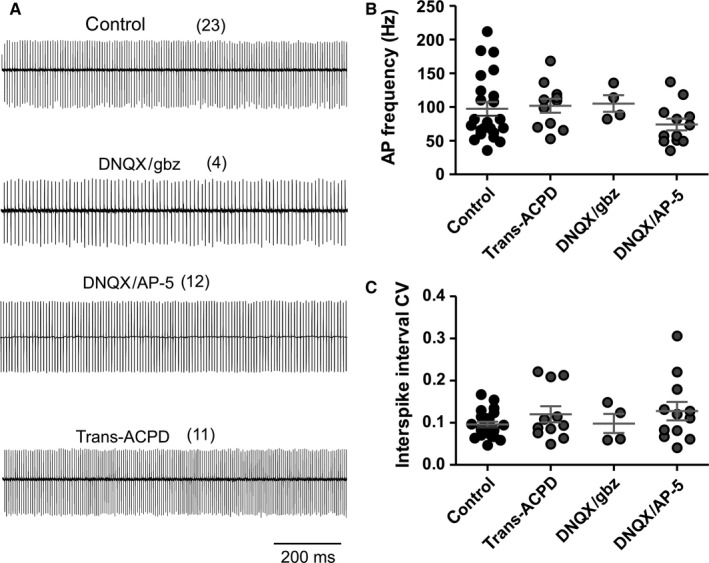
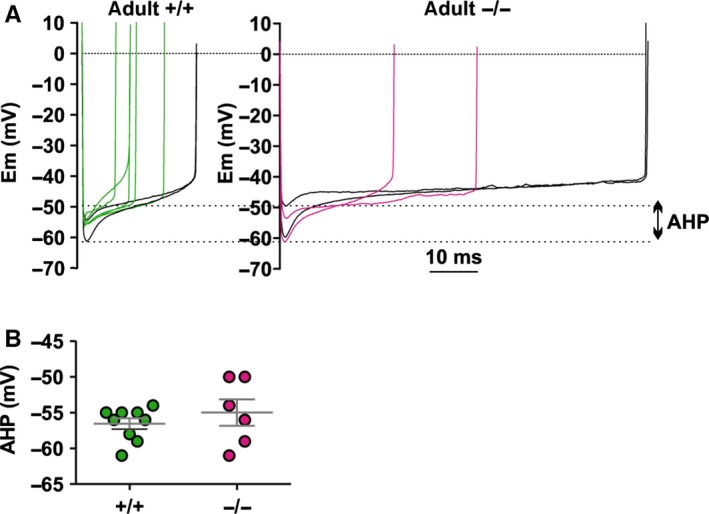
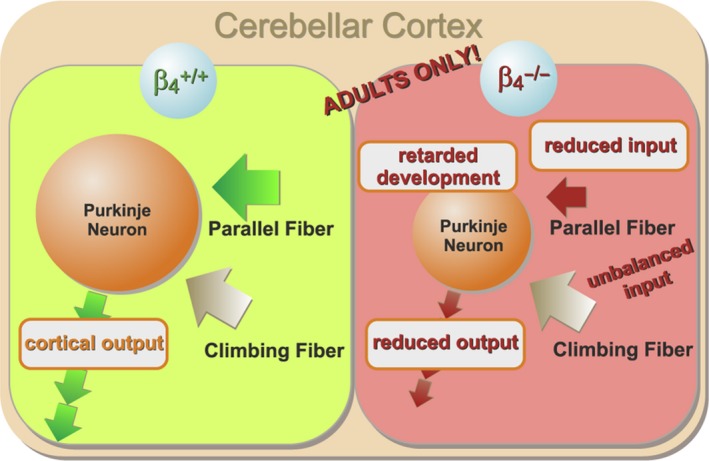
The auxiliary voltage-gated calcium channel subunit β4 supports targeting of calcium channels to the cell membrane, modulates ionic currents and promotes synaptic release in the central nervous system. β4 is abundant in cerebellum and its loss causes ataxia. However, the type of calcium channels and cerebellar functions affected by the loss of β4 are currently unknown. We therefore studied the structure and function of Purkinje cells in acute cerebellar slices of the β4 (-/-) ataxic (lethargic) mouse, finding that loss of β4 affected Purkinje cell input, morphology and pacemaker activity. In adult lethargic cerebellum evoked postsynaptic currents from parallel fibres were depressed, while paired-pulse facilitation and spontaneous synaptic currents were unaffected. Because climbing fibre input was spared, the parallel fibre/climbing fibre input ratio was reduced. The dendritic arbor of adult lethargic Purkinje cells displayed fewer and shorter dendrites, but a normal spine density. Accordingly, the width of the molecular and granular layers was reduced. These defects recapitulate the impaired cerebellar maturation observed upon Cav 2.1 ataxic mutations. However, unlike Cav 2.1 mutations, lethargic Purkinje cells also displayed a striking decrease in pacemaker firing frequency, without loss of firing regularity. All these deficiencies appear in late development, indicating the importance of β4 for the normal differentiation and function of mature Purkinje cells networks. The observed reduction of the parallel fibre input, the altered parallel fibre/climbing fibre ratio and the reduced Purkinje cell output can contribute to the severe motor impairment caused by the loss of the calcium channel β4 subunit in lethargic mice.
Keywords: ataxia; cerebellar cortex; development; electrophysiology; neuron.
© 2016 The Authors. European Journal of Neuroscience published by Federation of European Neuroscience Societies and John Wiley & Sons Ltd.
Figures
References
-
- Altman, J. (1972) Postnatal development of the cerebellar cortex in the rat. I. The external germinal layer and the transitional molecular layer. J. Comp. Neurol., 145, 353–397. - PubMed
-
- Barclay, J. , Balaguero, N. , Mione, M. , Ackerman, S.L. , Letts, V.A. , Brodbeck, J. , Canti, C. , Meir, A. et al (2001) Ducky mouse phenotype of epilepsy and ataxia is associated with mutations in the Cacna2d2 gene and decreased calcium channel current in cerebellar Purkinje cells. J. Neurosci., 21, 6095–6104. - PMC - PubMed
-
- Brodbeck, J. , Davies, A. , Courtney, J.M. , Meir, A. , Balaguero, N. , Canti, C. , Moss, F.J. , Page, K.M. et al (2002) The ducky mutation in Cacna2d2 results in altered Purkinje cell morphology and is associated with the expression of a truncated alpha 2 delta‐2 protein with abnormal function. J. Biol. Chem., 277, 7684–7693. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
