

D-Serine and Glycine Differentially Control Neurotransmission during Visual Cortex Critical Period
- PMID: 27003418
- PMCID: PMC4803205
- DOI: 10.1371/journal.pone.0151233
D-Serine and Glycine Differentially Control Neurotransmission during Visual Cortex Critical Period
Abstract
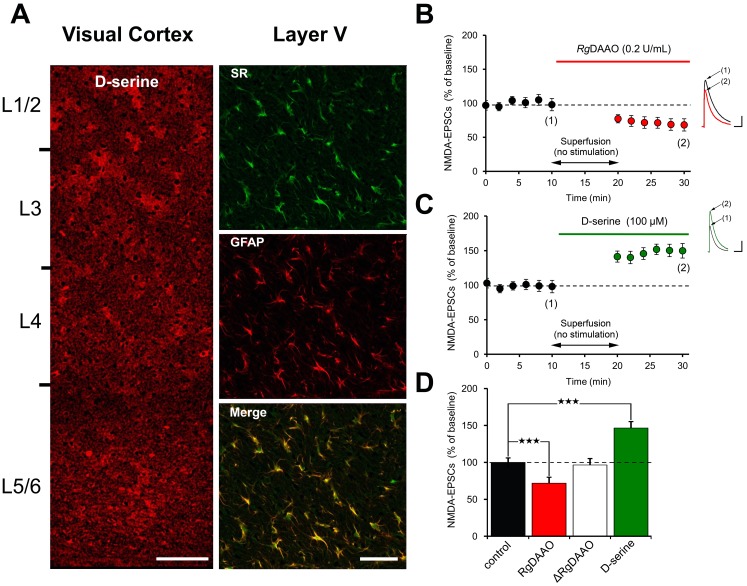
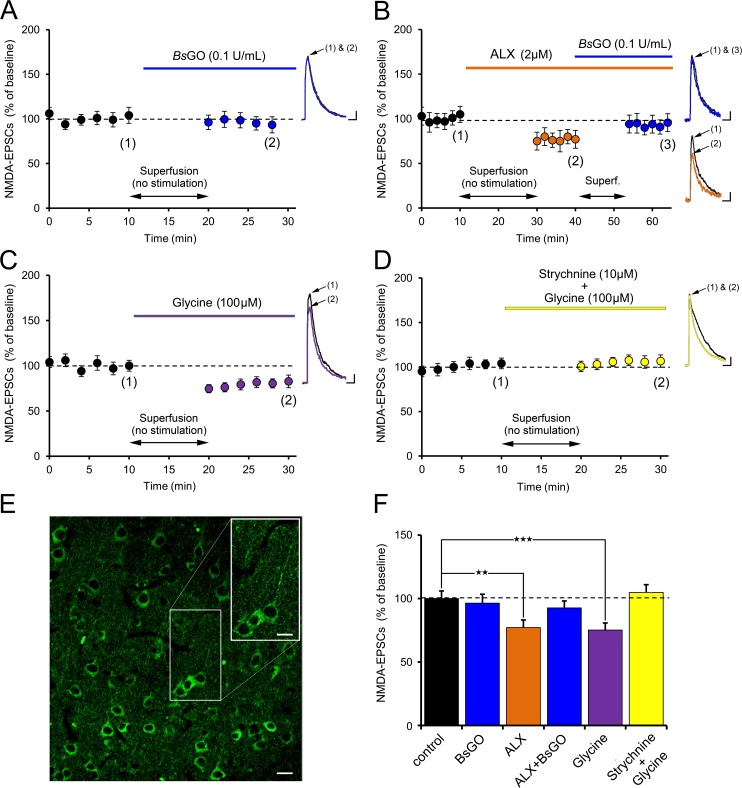
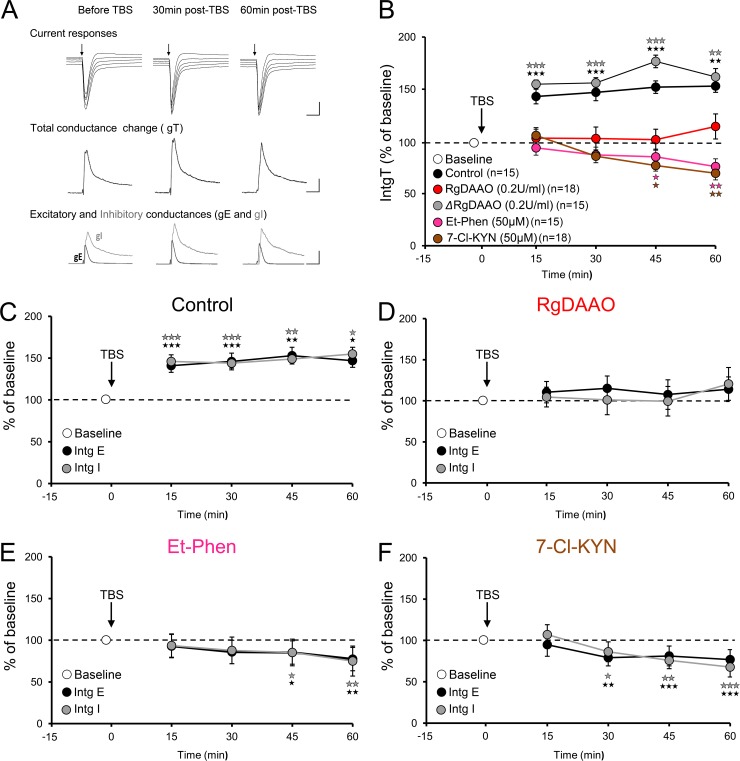
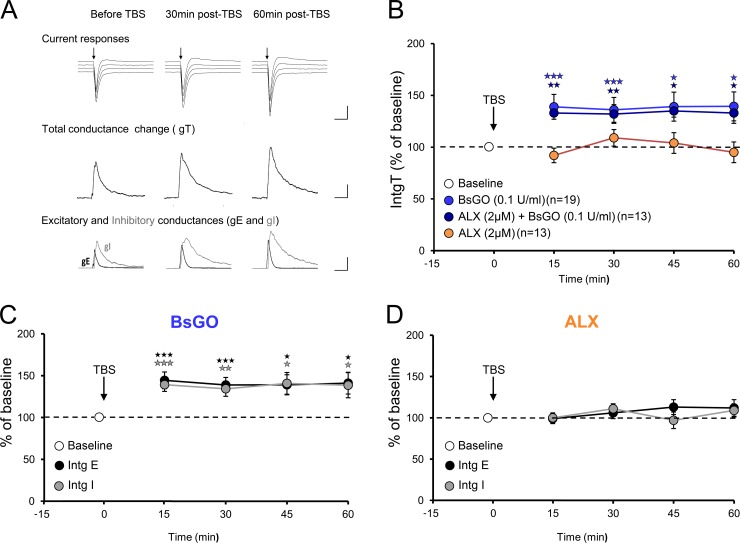
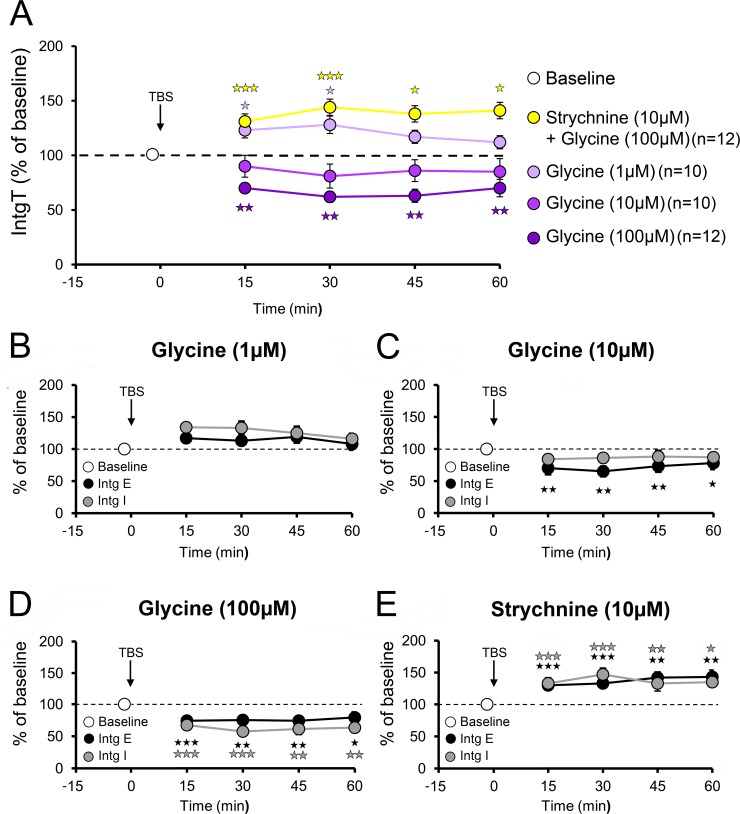
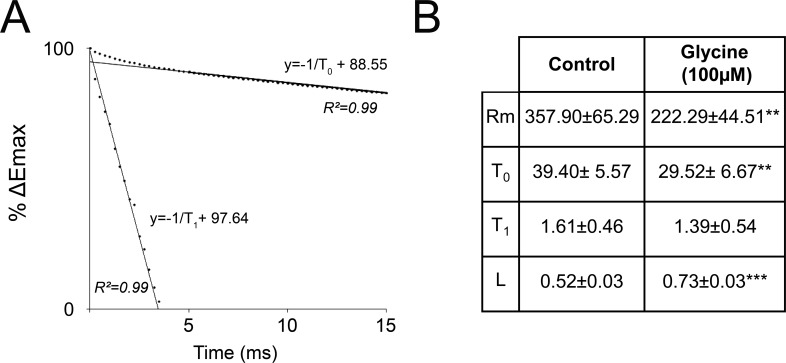
N-methyl-D-aspartate receptors (NMDARs) play a central role in synaptic plasticity. Their activation requires the binding of both glutamate and d-serine or glycine as co-agonist. The prevalence of either co-agonist on NMDA-receptor function differs between brain regions and remains undetermined in the visual cortex (VC) at the critical period of postnatal development. Here, we therefore investigated the regulatory role that d-serine and/or glycine may exert on NMDARs function and on synaptic plasticity in the rat VC layer 5 pyramidal neurons of young rats. Using selective enzymatic depletion of d-serine or glycine, we demonstrate that d-serine and not glycine is the endogenous co-agonist of synaptic NMDARs required for the induction and expression of Long Term Potentiation (LTP) at both excitatory and inhibitory synapses. Glycine on the other hand is not involved in synaptic efficacy per se but regulates excitatory and inhibitory neurotransmission by activating strychnine-sensitive glycine receptors, then producing a shunting inhibition that controls neuronal gain and results in a depression of synaptic inputs at the somatic level after dendritic integration. In conclusion, we describe for the first time that in the VC both D-serine and glycine differentially regulate somatic depolarization through the activation of distinct synaptic and extrasynaptic receptors.
Conflict of interest statement
Figures
References
-
- Kleckner NW, Dingledine R (1988) Requirement for glycine in activation of NMDA-receptors expressed in Xenopus oocytes. Science 241: 835–837. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
