

Decoding and Reconstructing the Focus of Spatial Attention from the Topography of Alpha-band Oscillations
- PMID: 27003790
- PMCID: PMC5074376
- DOI: 10.1162/jocn_a_00955
Decoding and Reconstructing the Focus of Spatial Attention from the Topography of Alpha-band Oscillations
Abstract
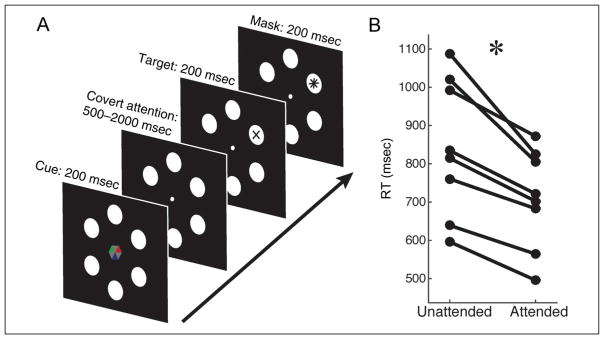
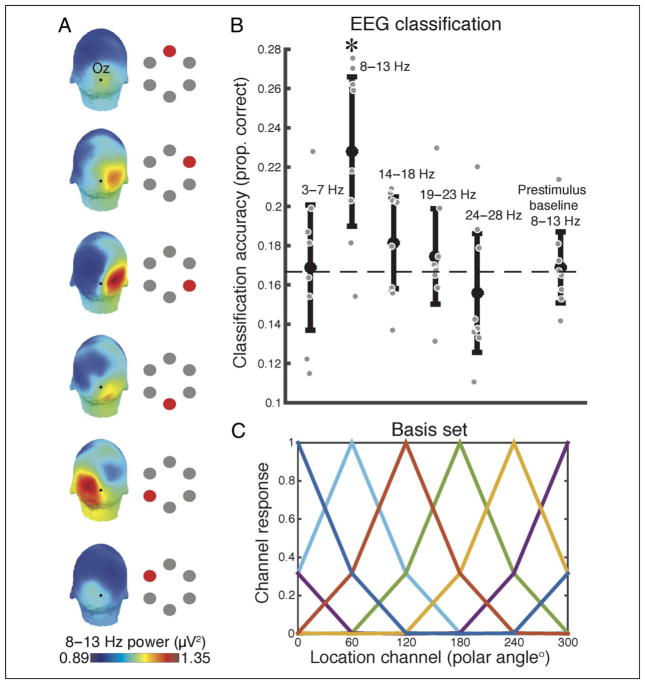
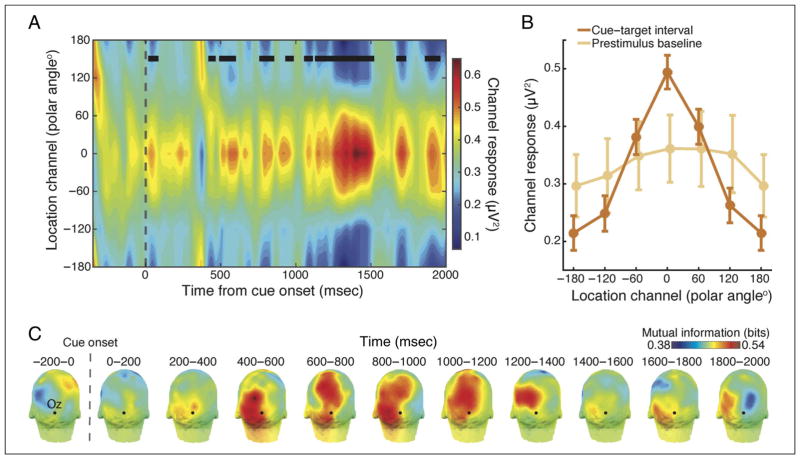
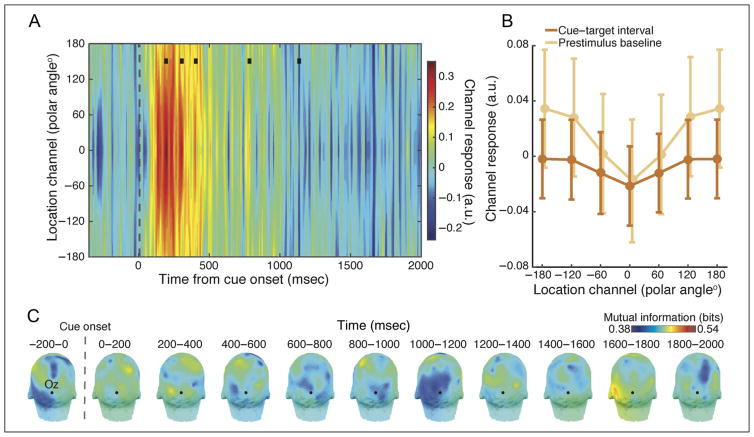
Many aspects of perception and cognition are supported by activity in neural populations that are tuned to different stimulus features (e.g., orientation, spatial location, color). Goal-directed behavior, such as sustained attention, requires a mechanism for the selective prioritization of contextually appropriate representations. A candidate mechanism of sustained spatial attention is neural activity in the alpha band (8-13 Hz), whose power in the human EEG covaries with the focus of covert attention. Here, we applied an inverted encoding model to assess whether spatially selective neural responses could be recovered from the topography of alpha-band oscillations during spatial attention. Participants were cued to covertly attend to one of six spatial locations arranged concentrically around fixation while EEG was recorded. A linear classifier applied to EEG data during sustained attention demonstrated successful classification of the attended location from the topography of alpha power, although not from other frequency bands. We next sought to reconstruct the focus of spatial attention over time by applying inverted encoding models to the topography of alpha power and phase. Alpha power, but not phase, allowed for robust reconstructions of the specific attended location beginning around 450 msec postcue, an onset earlier than previous reports. These results demonstrate that posterior alpha-band oscillations can be used to track activity in feature-selective neural populations with high temporal precision during the deployment of covert spatial attention.
Figures
References
-
- Bahramisharif A, van Gerven M, Heskes T, Jensen O. Covert attention allows for continuous control of brain-computer interfaces. European Journal of Neuroscience. 2010;31:1501–1508. - PubMed
-
- Bonnefond M, Jensen O. Alpha oscillations serve to protect working memory maintenance against anticipated distracters. Current Biology. 2012;22:1969–1974. - PubMed
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources