

Prohibitin-induced, obesity-associated insulin resistance and accompanying low-grade inflammation causes NASH and HCC
- PMID: 27005704
- PMCID: PMC4804274
- DOI: 10.1038/srep23608
Prohibitin-induced, obesity-associated insulin resistance and accompanying low-grade inflammation causes NASH and HCC
Abstract
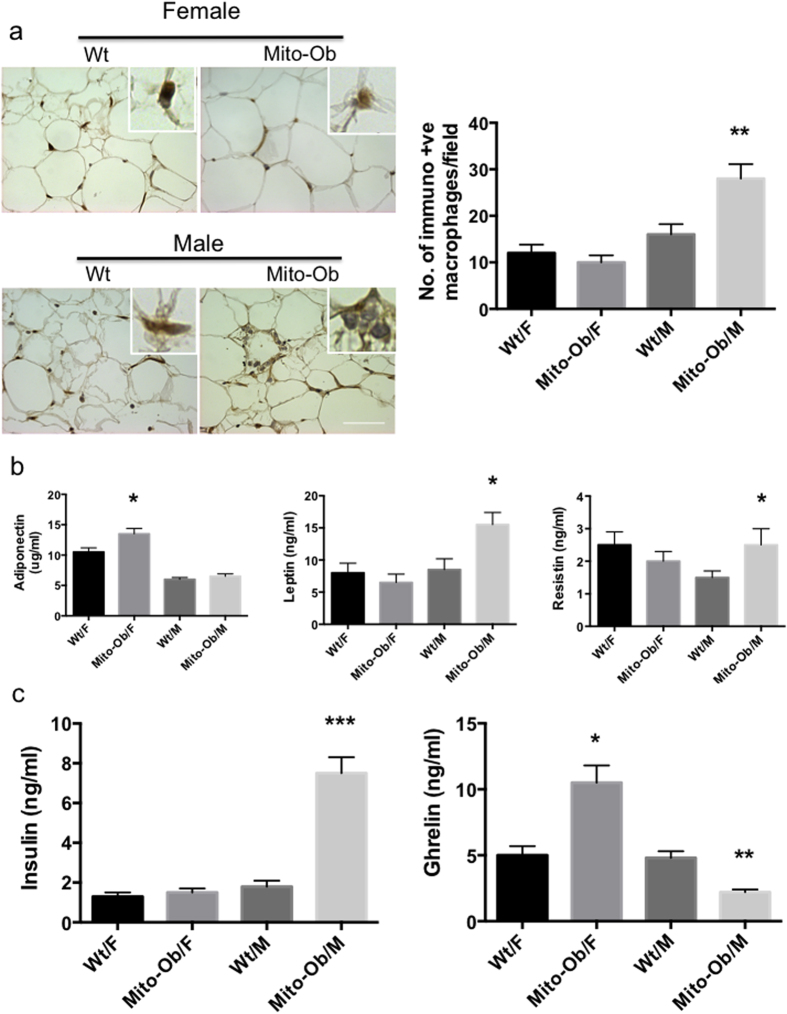
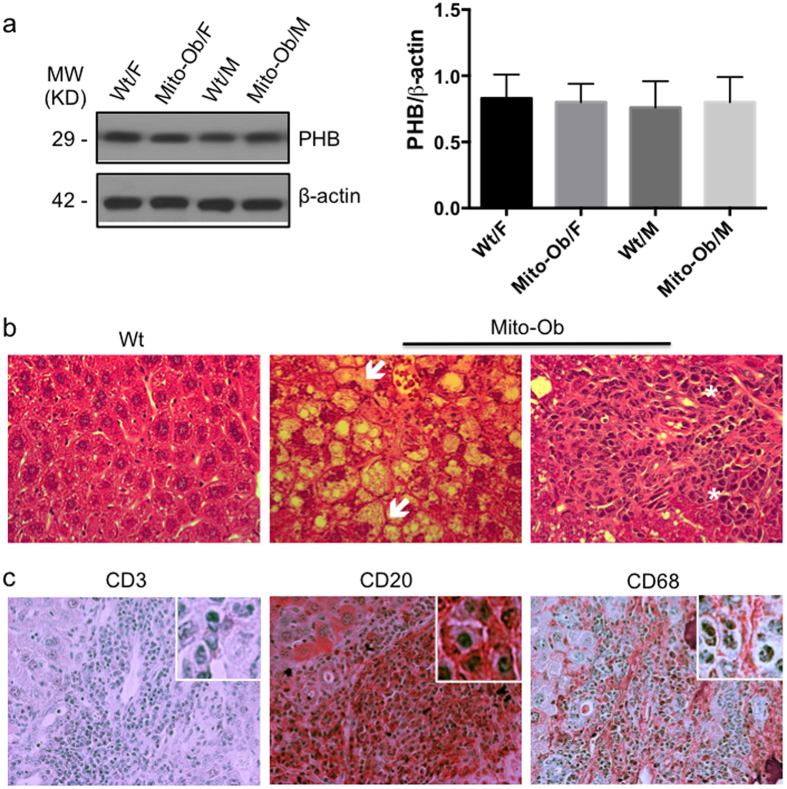
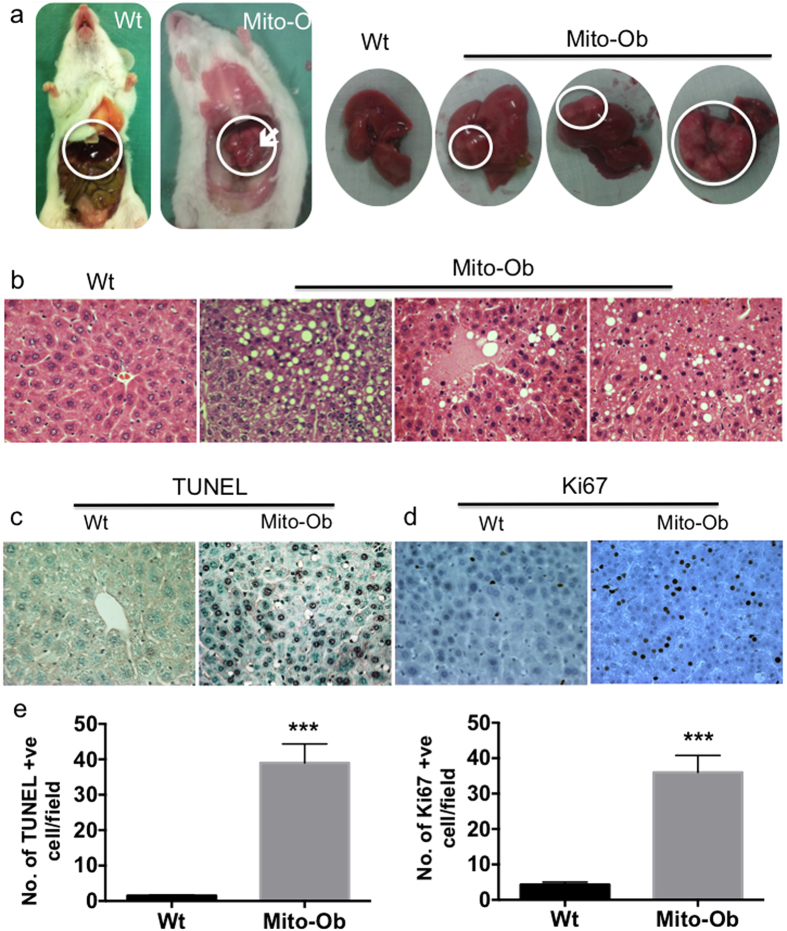
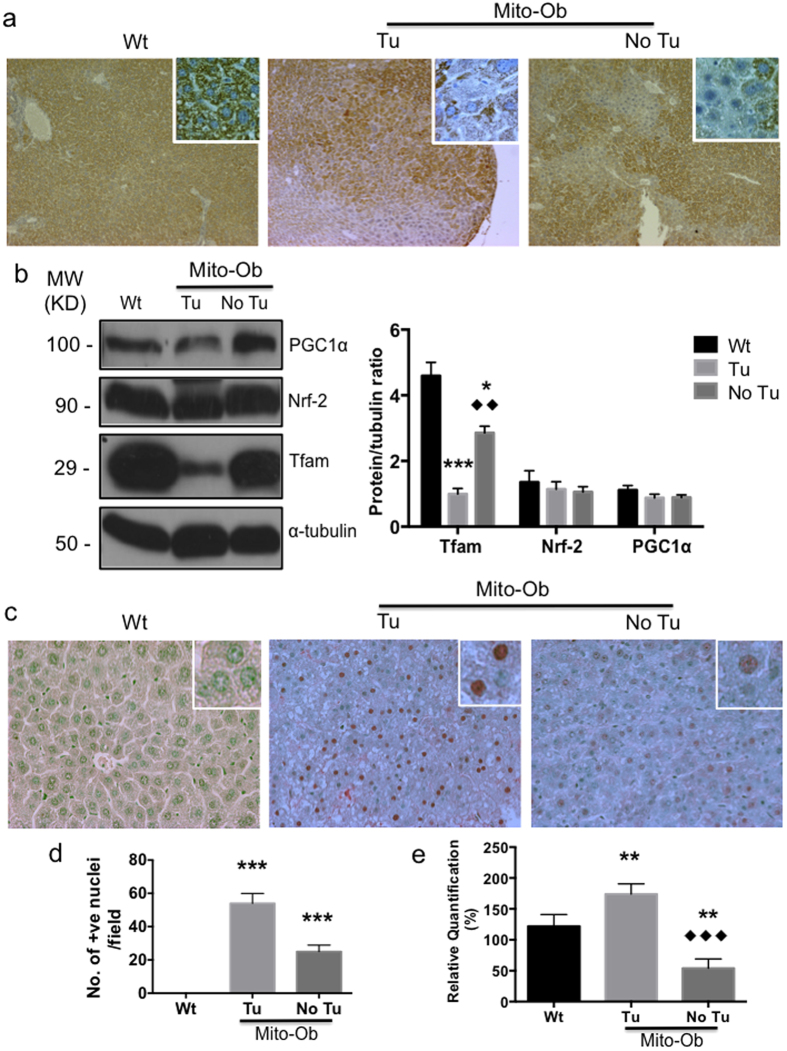
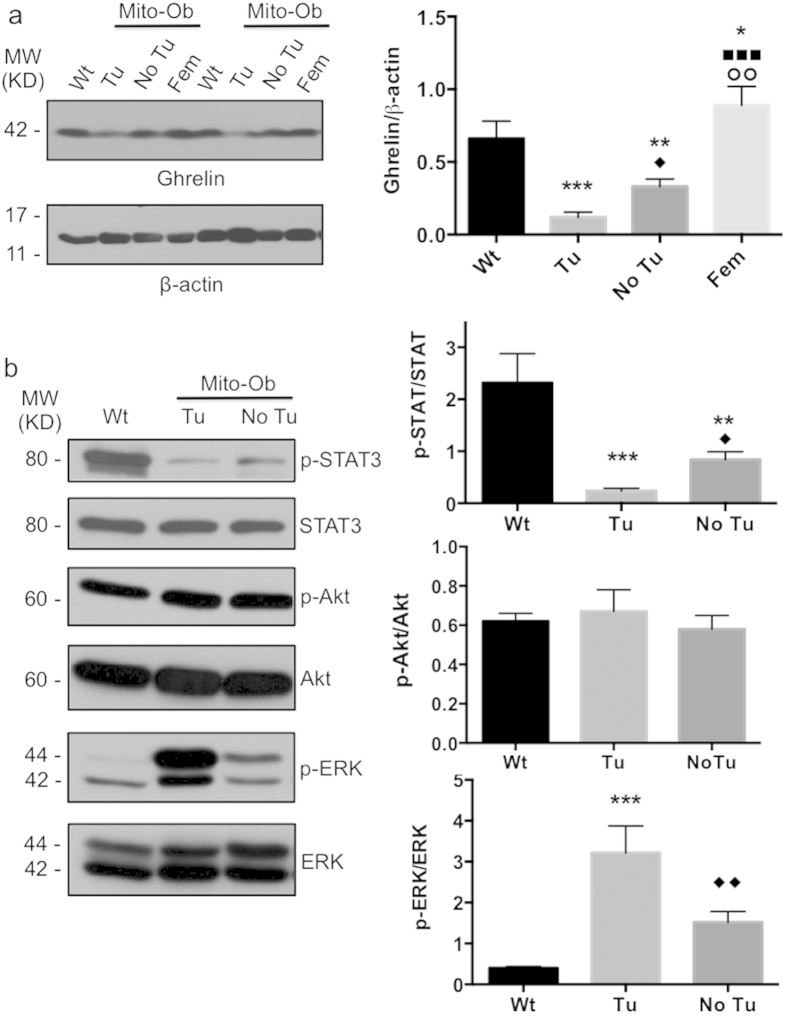
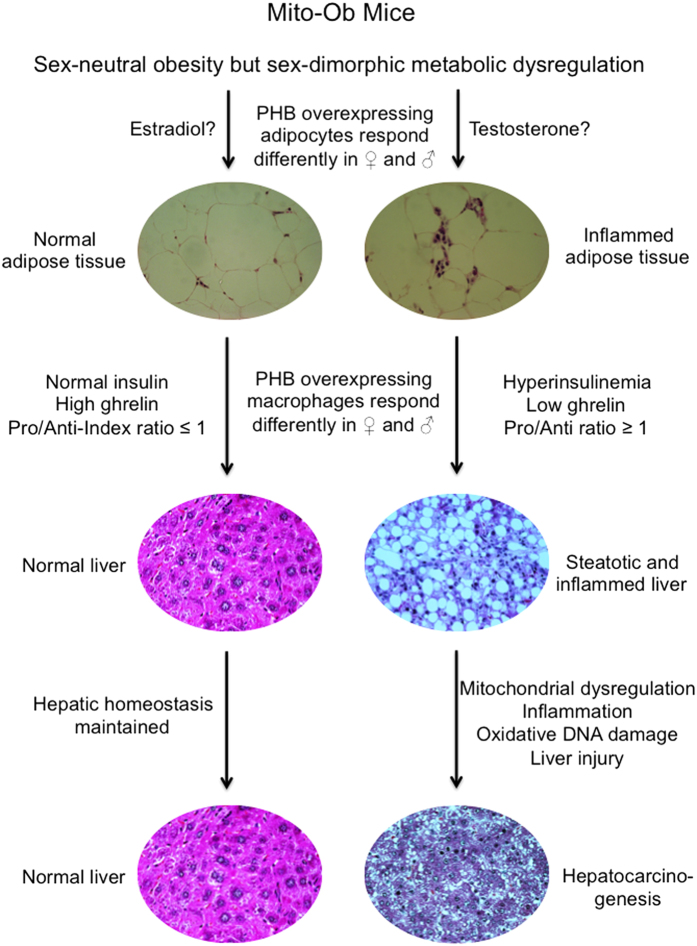
Obesity increases the risk for nonalcoholic steatohepatitis (NASH) and hepatocarcinogenesis. However, the underlying mechanisms involved in the disease process remain unclear. Recently, we have developed a transgenic obese mouse model (Mito-Ob) by prohibitin mediated mitochondrial remodeling in adipocytes. The Mito-Ob mice develop obesity in a sex-neutral manner, but obesity-associated adipose inflammation and metabolic dysregulation in a male sex-specific manner. Here we report that with aging, the male Mito-Ob mice spontaneously develop obesity-linked NASH and hepatocellular carcinoma (HCC). In contrast, the female Mito-Ob mice maintained normal glucose and insulin levels and did not develop NASH and HCC. The anti-inflammatory peptide ghrelin was significantly upregulated in the female mice and down regulated in the male mice compared with respective control mice. In addition, a reduction in the markers of mitochondrial content and function was found in the liver of male Mito-Ob mice with NASH/HCC development. We found that ERK1/2 signaling was significantly upregulated whereas STAT3 signaling was significantly down regulated in the tumors from Mito-Ob mice. These data provide a proof-of-concept that the metabolic and inflammatory status of the adipose tissue and their interplay at the systemic and hepatic level play a central role in the pathogenesis of obesity-linked NASH and HCC.
Figures
References
-
- El-Serag H. B. Hepatocellular carcinoma. N Engl J Med. 365, 118–127 (2011). - PubMed
-
- Tilg H. & Moschen A. R. Mechanisms behind the link between obesity and gastrointestinal cancers. Best Pract Res Clin Gastroenterol. 28, 599–610 (2014). - PubMed
-
- Calle E. E. et al. Overweight, obesity and mortality from cancer in a prospective studied cohort of U.S. adults. N Engl J Med 348, 1625–1638 (2003). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
