

The key role of CYC2 during meiosis in Tetrahymena thermophila
- PMID: 27008457
- PMCID: PMC4818844
- DOI: 10.1007/s13238-016-0254-9
The key role of CYC2 during meiosis in Tetrahymena thermophila
Erratum in
-
Correction to: The key role of CYC2 during meiosis in Tetrahymena Thermophila.Protein Cell. 2018 Nov;9(11):981. doi: 10.1007/s13238-018-0562-3. Protein Cell. 2018. PMID: 29951792 Free PMC article.
Abstract
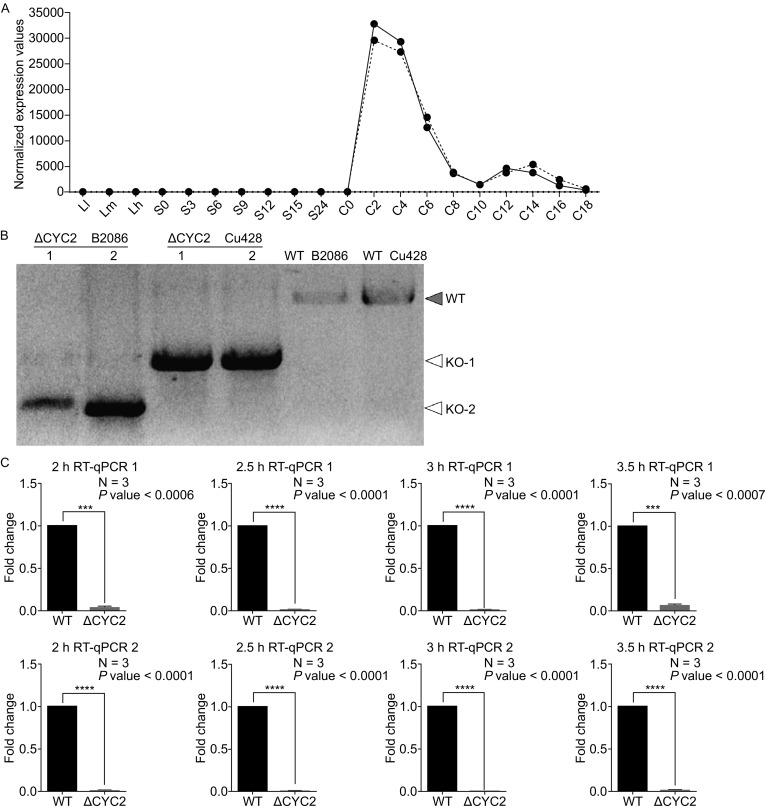
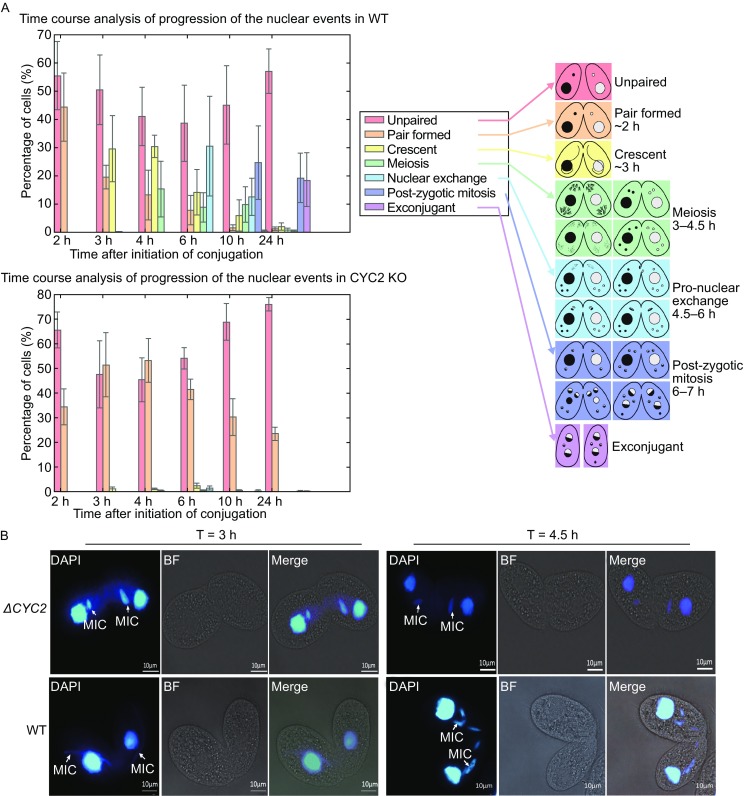
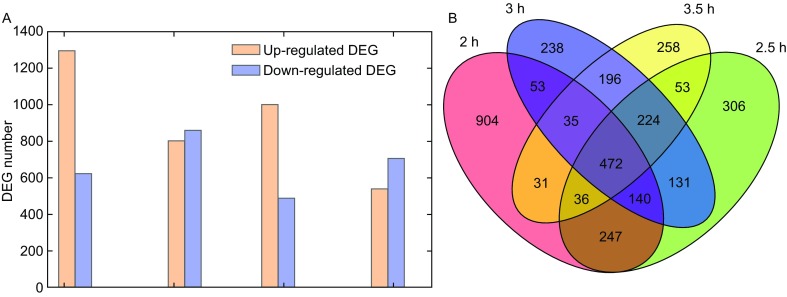
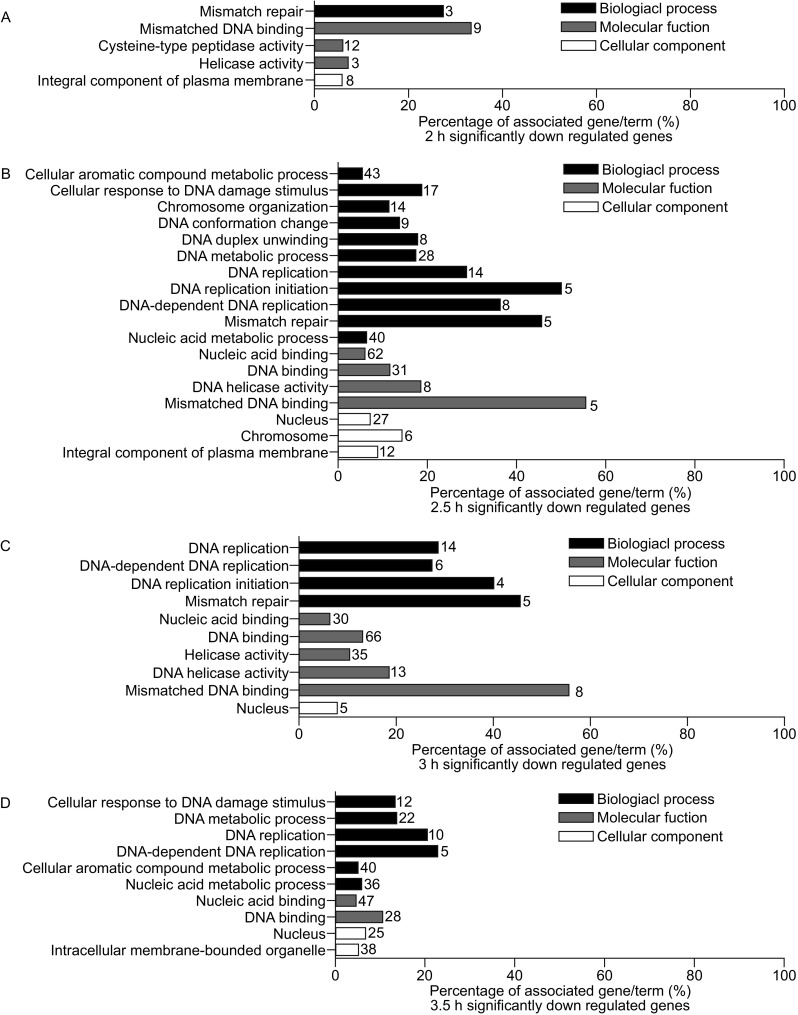
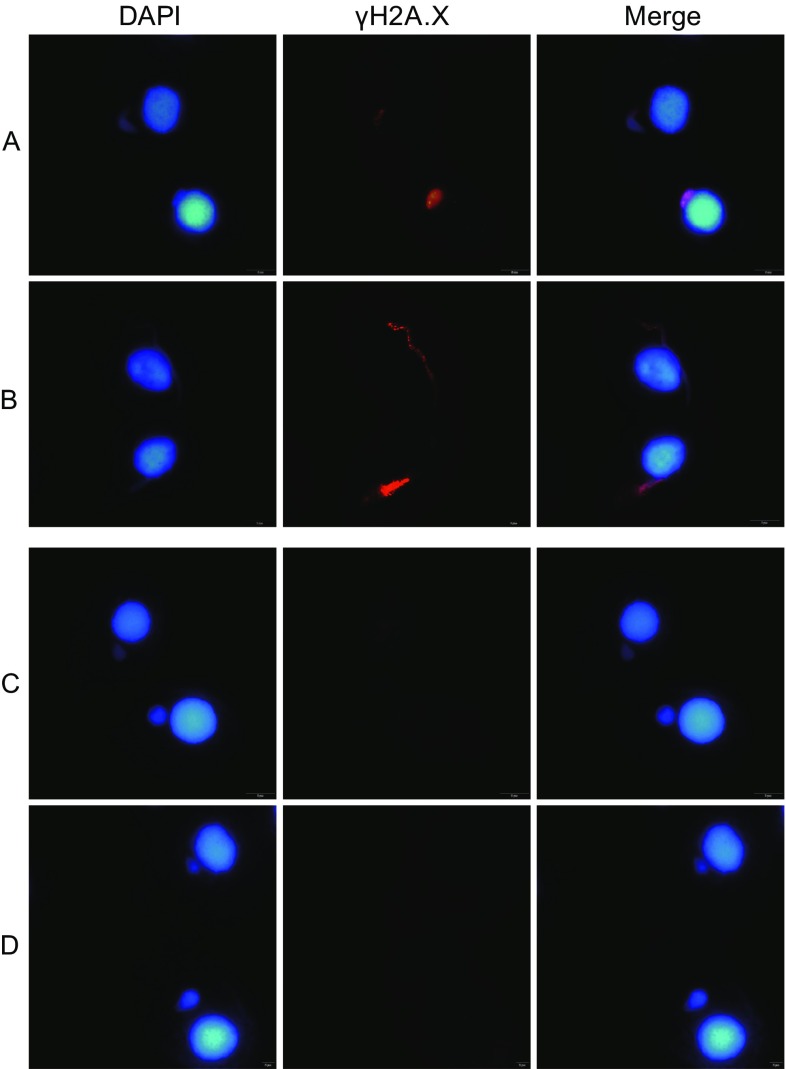
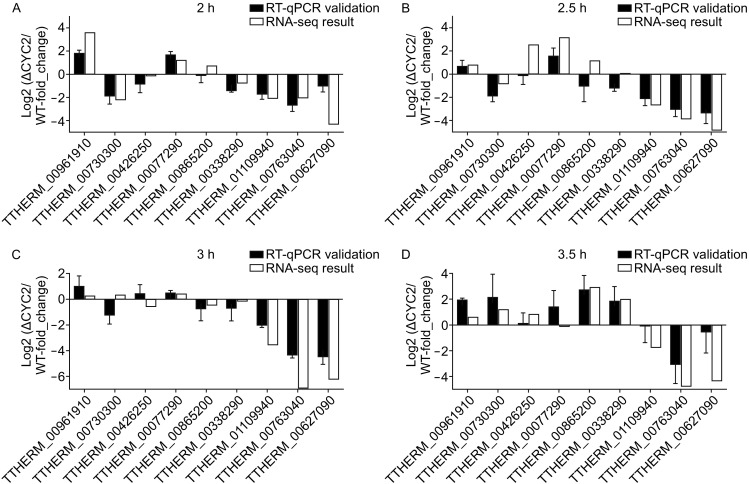
Meiotic recombination is carried out through a specialized pathway for the formation and repair of DNA double-strand breaks (DSBs) made by the Spo11 protein. The present study shed light on the functional role of cyclin, CYC2, in Tetrahymena thermophila which has transcriptionally high expression level during meiosis process. Knocking out the CYC2 gene results in arrest of meiotic conjugation process at 2.5-3.5 h after conjugation initiation, before the meiosis division starts, and in company with the absence of DSBs. To investigate the underlying mechanism of this phenomenon, a complete transcriptome profile was performed between wild-type strain and CYC2 knock-out strain. Functional analysis of RNA-Seq results identifies related differentially expressed genes (DEGs) including SPO11 and these DEGs are enriched in DNA repair/mismatch repair (MMR) terms in homologous recombination (HR), which indicates that CYC2 could play a crucial role in meiosis by regulating SPO11 and participating in HR.
Keywords: RNA-Seq; Tetrahymena thermophila; cyclin; homologous recombination; meiosis.
Figures
References
-
- Bindea G, Mlecnik B, Hackl H, Charoentong P, Tosolini M, Kirilovsky A, Fridman WH, Pages F, Trajanoski Z, Galon J. ClueGO: a cytoscape plug-into decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics. 2009;25:1091–1093. doi: 10.1093/bioinformatics/btp101. - DOI - PMC - PubMed
-
- Collins K. Perspectives on the ciliated protozoan Tetrahymena thermophila. Method Cell Biol. 2012;109:3–7. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
