

Biophysical constraints on lateral inhibition in the olfactory bulb
- PMID: 27009162
- PMCID: PMC4922614
- DOI: 10.1152/jn.00671.2015
Biophysical constraints on lateral inhibition in the olfactory bulb
Abstract
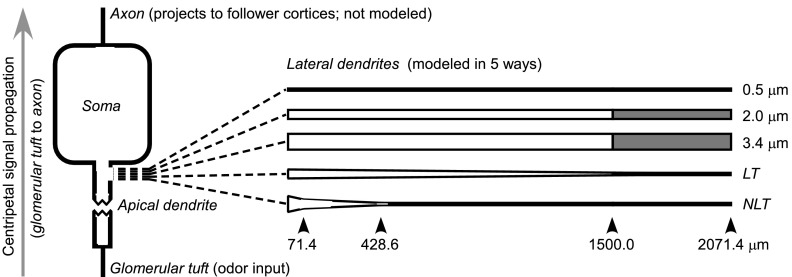
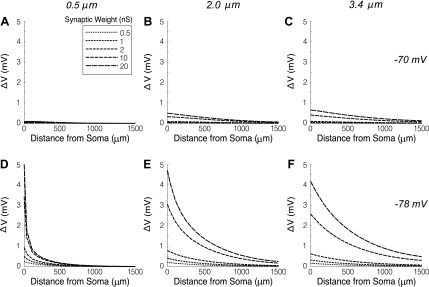
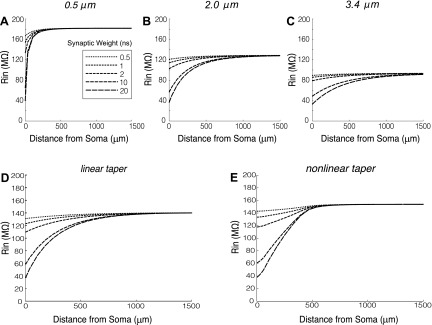
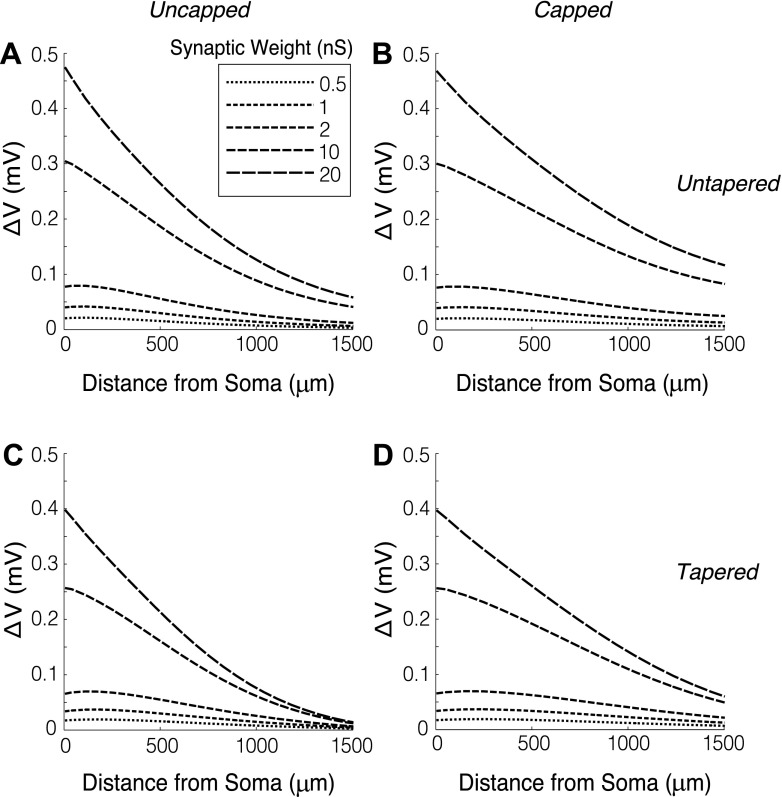
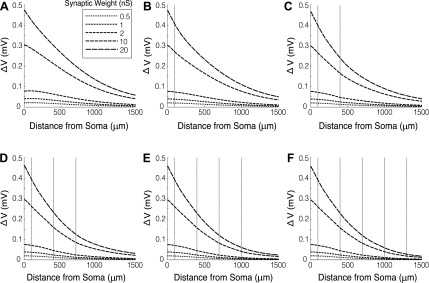
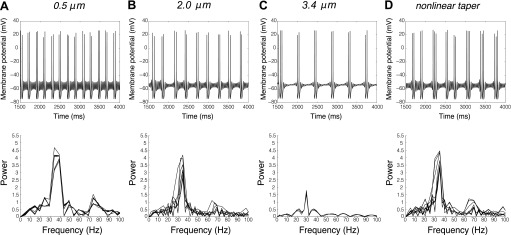
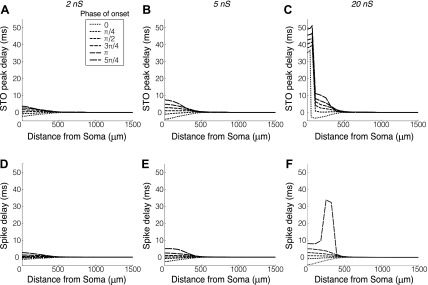
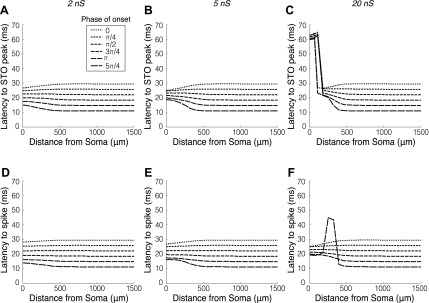
The mitral cells (MCs) of the mammalian olfactory bulb (OB) constitute one of two populations of principal neurons (along with middle/deep tufted cells) that integrate afferent olfactory information with top-down inputs and intrinsic learning and deliver output to downstream olfactory areas. MC activity is regulated in part by inhibition from granule cells, which form reciprocal synapses with MCs along the extents of their lateral dendrites. However, with MC lateral dendrites reaching over 1.5 mm in length in rats, the roles of distal inhibitory synapses pose a quandary. Here, we systematically vary the properties of a MC model to assess the capacity of inhibitory synaptic inputs on lateral dendrites to influence afferent information flow through MCs. Simulations using passivized models with varying dendritic morphologies and synaptic properties demonstrated that, even with unrealistically favorable parameters, passive propagation fails to convey effective inhibitory signals to the soma from distal sources. Additional simulations using an active model exhibiting action potentials, subthreshold oscillations, and a dendritic morphology closely matched to experimental values further confirmed that distal synaptic inputs along the lateral dendrite could not exert physiologically relevant effects on MC spike timing at the soma. Larger synaptic conductances representative of multiple simultaneous inputs were not sufficient to compensate for the decline in signal with distance. Reciprocal synapses on distal MC lateral dendrites may instead serve to maintain a common fast oscillatory clock across the OB by delaying spike propagation within the lateral dendrites themselves.
Keywords: cable theory; computational neuroscience; lateral dendrites; mitral cell; olfaction.
Copyright © 2016 the American Physiological Society.
Figures
Similar articles
-
Multiple modes of action potential initiation and propagation in mitral cell primary dendrite.J Neurophysiol. 2002 Nov;88(5):2755-64. doi: 10.1152/jn.00057.2002. J Neurophysiol. 2002. PMID: 12424310
-
Inhibition of backpropagating action potentials in mitral cell secondary dendrites.J Neurophysiol. 2002 Jul;88(1):64-85. doi: 10.1152/jn.2002.88.1.64. J Neurophysiol. 2002. PMID: 12091533
-
Organization of inhibition in the rat olfactory bulb external plexiform layer.J Neurophysiol. 1993 Jul;70(1):263-74. doi: 10.1152/jn.1993.70.1.263. J Neurophysiol. 1993. PMID: 8395579
-
Predicting brain organization with a computational model: 50-year perspective on lateral inhibition and oscillatory gating by dendrodendritic synapses.J Neurophysiol. 2020 Aug 1;124(2):375-387. doi: 10.1152/jn.00175.2020. Epub 2020 Jul 8. J Neurophysiol. 2020. PMID: 32639901 Free PMC article. Review.
-
Lateral inhibition in the olfactory bulb and in olfaction.Physiol Behav. 2002 Dec;77(4-5):607-12. doi: 10.1016/s0031-9384(02)00895-8. Physiol Behav. 2002. PMID: 12527007 Review.
Cited by
-
Reinforced Odor Representations in the Anterior Olfactory Nucleus Can Serve as Memory Traces for Conspecifics.eNeuro. 2025 Jul 28;12(7):ENEURO.0143-25.2025. doi: 10.1523/ENEURO.0143-25.2025. Print 2025 Jul. eNeuro. 2025. PMID: 40664512 Free PMC article.
-
Anatomical and Functional Connectivity at the Dendrodendritic Reciprocal Mitral Cell-Granule Cell Synapse: Impact on Recurrent and Lateral Inhibition.Front Neural Circuits. 2022 Jul 22;16:933201. doi: 10.3389/fncir.2022.933201. eCollection 2022. Front Neural Circuits. 2022. PMID: 35937203 Free PMC article.
-
Presynaptic NMDARs cooperate with local spikes toward GABA release from the reciprocal olfactory bulb granule cell spine.Elife. 2020 Nov 30;9:e63737. doi: 10.7554/eLife.63737. Elife. 2020. PMID: 33252329 Free PMC article.
-
Context-dependent odor learning requires the anterior olfactory nucleus.Behav Neurosci. 2020 Aug;134(4):332-343. doi: 10.1037/bne0000371. Epub 2020 May 7. Behav Neurosci. 2020. PMID: 32378908 Free PMC article.
-
A coupled-oscillator model of olfactory bulb gamma oscillations.PLoS Comput Biol. 2017 Nov 15;13(11):e1005760. doi: 10.1371/journal.pcbi.1005760. eCollection 2017 Nov. PLoS Comput Biol. 2017. PMID: 29140973 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous