

Prediction of the in planta Phakopsora pachyrhizi secretome and potential effector families
- PMID: 27010366
- PMCID: PMC6638266
- DOI: 10.1111/mpp.12405
Prediction of the in planta Phakopsora pachyrhizi secretome and potential effector families
Abstract
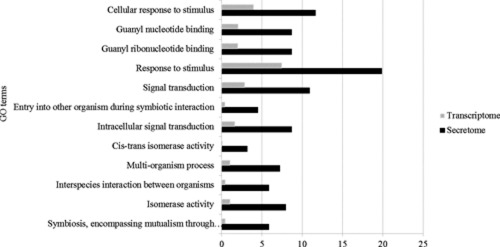
Asian soybean rust (ASR), caused by the obligate biotrophic fungus Phakopsora pachyrhizi, can cause losses greater than 80%. Despite its economic importance, there is no soybean cultivar with durable ASR resistance. In addition, the P. pachyrhizi genome is not yet available. However, the availability of other rust genomes, as well as the development of sample enrichment strategies and bioinformatics tools, has improved our knowledge of the ASR secretome and its potential effectors. In this context, we used a combination of laser capture microdissection (LCM), RNAseq and a bioinformatics pipeline to identify a total of 36 350 P. pachyrhizi contigs expressed in planta and a predicted secretome of 851 proteins. Some of the predicted secreted proteins had characteristics of candidate effectors: small size, cysteine rich, do not contain PFAM domains (except those associated with pathogenicity) and strongly expressed in planta. A comparative analysis of the predicted secreted proteins present in Pucciniales species identified new members of soybean rust and new Pucciniales- or P. pachyrhizi-specific families (tribes). Members of some families were strongly up-regulated during early infection, starting with initial infection through haustorium formation. Effector candidates selected from two of these families were able to suppress immunity in transient assays, and were localized in the plant cytoplasm and nuclei. These experiments support our bioinformatics predictions and show that these families contain members that have functions consistent with P. pachyrhizi effectors.
Keywords: ETI suppression; LCM; P. pachyrhizi; effectors; secretome; soybean rust; transcriptome.
© 2016 BSPP AND JOHN WILEY & SONS LTD.
Figures




References
-
- Aimanianda, V. , Bayry, J. , Bozza, S. , Kniemeyer, O. , Perruccio, K. , Elluru, S.R. , Clavaud, C. , Paris, S. , Brakhage, A.A. , Kaveri, S.V. , Romani, L. and Latgé, J.P. (2009) Surface hydrophobin prevents immune recognition of airborne fungal spores. Nature, 460, 1117–1121. - PubMed
-
- Aime, M.C. , Matheny, P.B. , Henk, D.A. , Frieders, E.M. , Nilsson, R.H. , Piepenbring, M. , McLaughlin, D.J. , Szabo, L.J. , Begerow, D. , Sampaio, J.P. , Bauer, R. , Weiß, M. , Oberwinkler, F. and Hibbett, D. (2006) An overview of the higher level classification of Pucciniomycotina based on combined analyses of nuclear large and small subunit rDNA sequences. Mycologia, 98, 896–905. - PubMed
-
- Bailey, T.L. and Elkan, C. (1994) Fitting a mixture model by expectation maximization to discover motifs in biopolymers. Proc. Int. Conf. Intell. Syst. Mol. Biol. 2, 28–36. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
