

Mechanisms and consequences of aneuploidy and chromosome instability in the aging brain
- PMID: 27013377
- PMCID: PMC5490080
- DOI: 10.1016/j.mad.2016.03.007
Mechanisms and consequences of aneuploidy and chromosome instability in the aging brain
Abstract
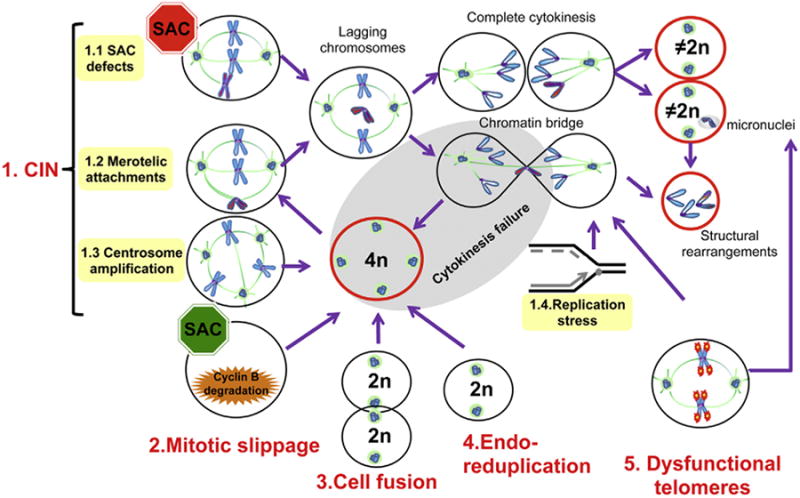
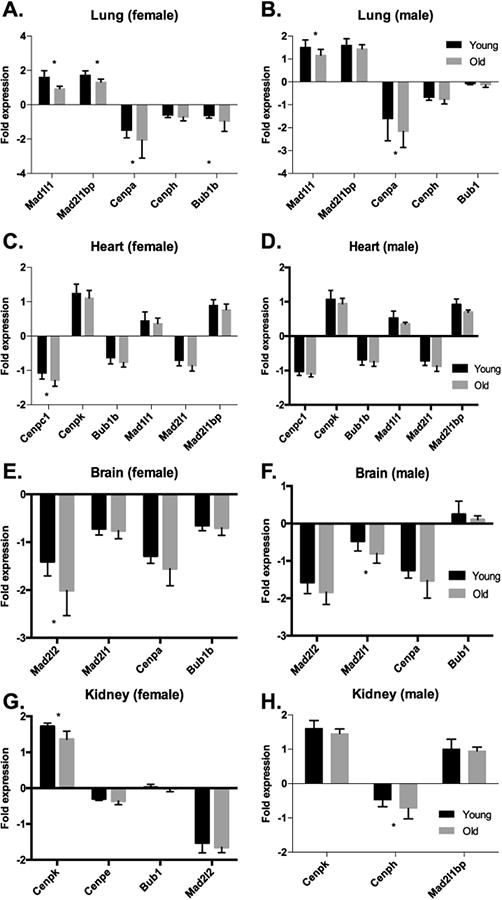
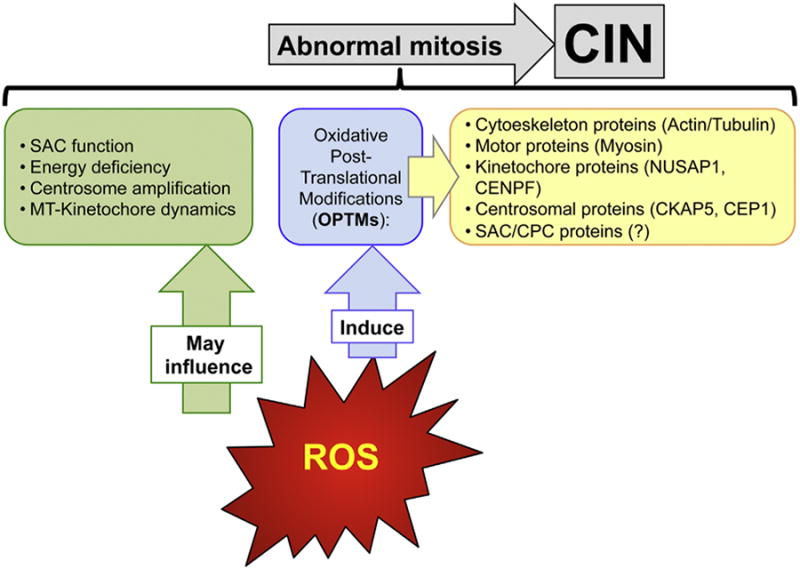
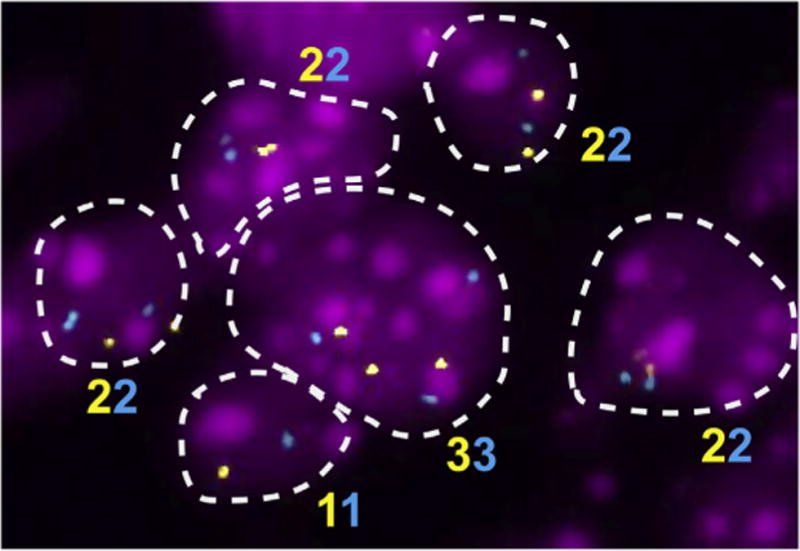
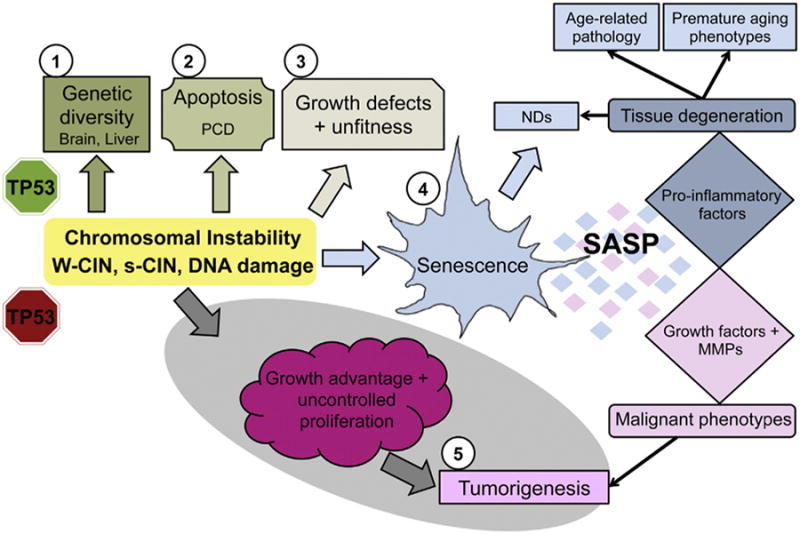
Aneuploidy and polyploidy are a form of Genomic Instability (GIN) known as Chromosomal Instability (CIN) characterized by sporadic abnormalities in chromosome copy numbers. Aneuploidy is commonly linked to pathological states. It is a hallmark of spontaneous abortions and birth defects and it is observed virtually in every human tumor, therefore being generally regarded as detrimental for the development or the maturation of tissues under physiological conditions. Polyploidy however, occurs as part of normal physiological processes during maturation and differentiation of some mammalian cell types. Surprisingly, high levels of aneuploidy are present in the brain, and their frequency increases with age suggesting that the brain is able to maintain its functionality in the presence of high levels of mosaic aneuploidy. Because somatic aneuploidy with age can reach exceptionally high levels, it is likely to have long-term adverse effects in this organ. We describe the mechanisms accountable for an abnormal DNA content with a particular emphasis on the CNS where cell division is limited. Next, we briefly summarize the types of GIN known to date and discuss how they interconnect with CIN. Lastly we highlight how several forms of CIN may contribute to genetic variation, tissue degeneration and disease in the CNS.
Keywords: Aneuploidy; Brain; DNA damage; Genomic instability; Polyploidy; Tissue degeneration; Whole chromosome instability (W-CIN).
Copyright © 2016. Published by Elsevier B.V.
Figures
References
-
- Alvarez-Dolado M, Pardal R, Garcia-Verdugo JM, Fike JR, Lee HO, Pfeffer K, Lois C, Morrison SJ, Alvarez-Buylla A. Fusion of bone-marrow-derived cells with Purkinje neurons, cardiomyocytes and hepatocytes. Nature. 2003;425:968–973. - PubMed
-
- Ambros M, Rumpler S, Luegmayr A, Hattinger CM, Strehl S, Kovar H, Gadner H, Ambros PF. Neuroblastoma cells can actively eliminate supernumerary MYCN gene copies by micronucleus formation—sign of tumourcell revertance? Eur J Cancer. 1997;33(12):2043–2049. - PubMed
-
- Arocena DG, Iwahashi CK, Won N, Beilina A, Ludwig AL, Tassone F, Schwartz PH, Hagerman PJ. Induction of inclusion formation and disruption of lamin A/C structure by premutation CGG-repeat RNA in human cultured neural cells. Hum Mol Genet. 2005;14:3661–3671. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
