

Nuclear envelope rupture and repair during cancer cell migration
- PMID: 27013428
- PMCID: PMC4833568
- DOI: 10.1126/science.aad7297
Nuclear envelope rupture and repair during cancer cell migration
Abstract
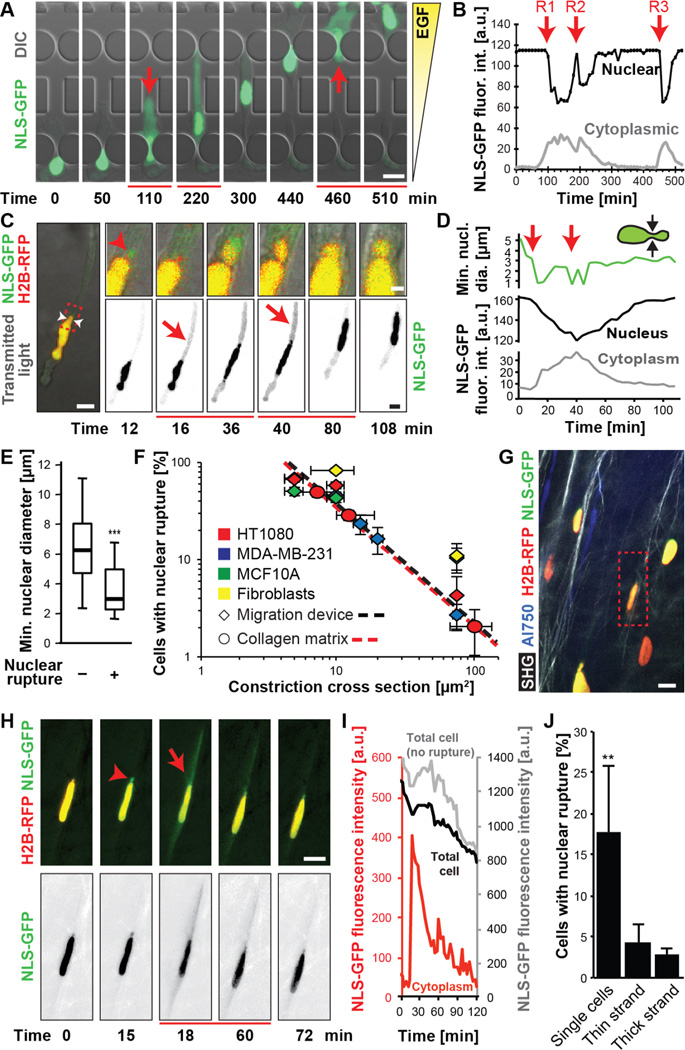
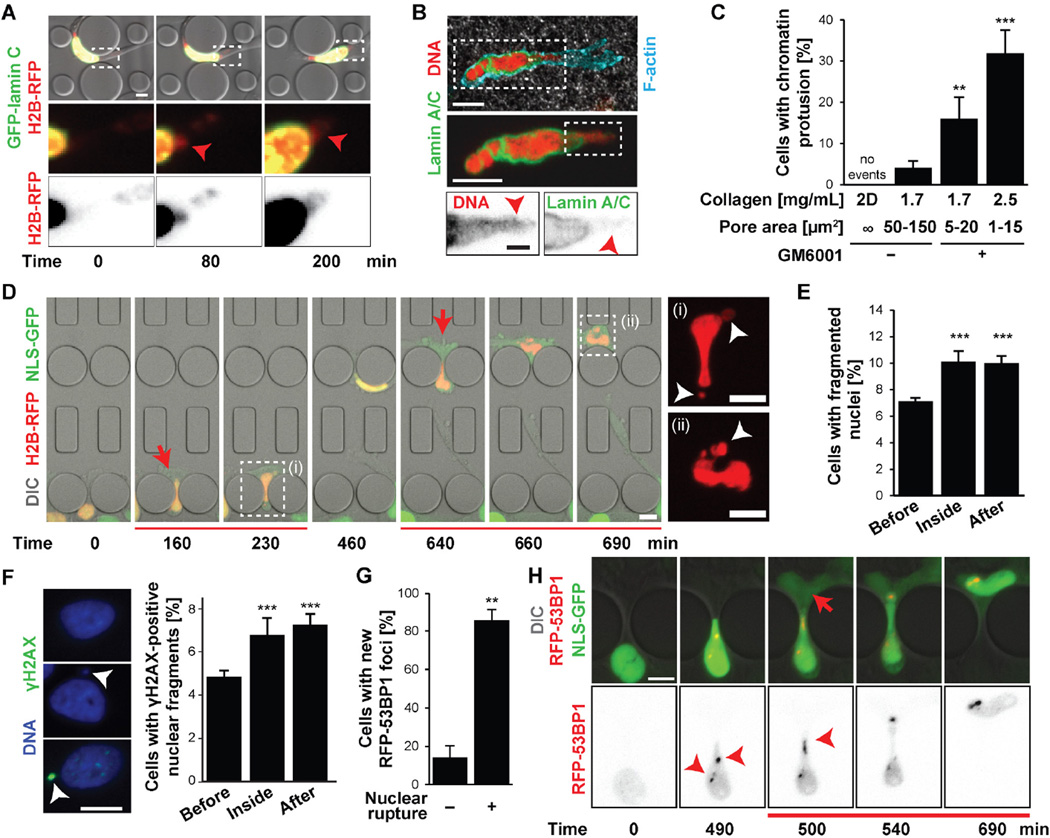
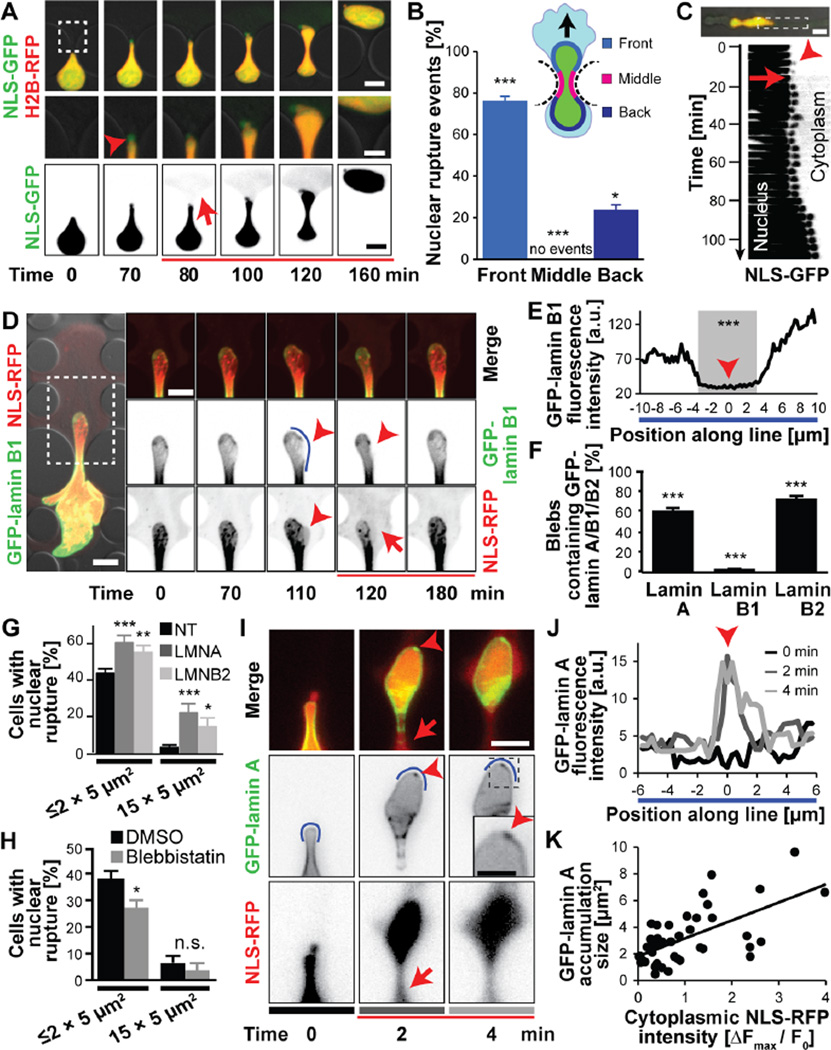
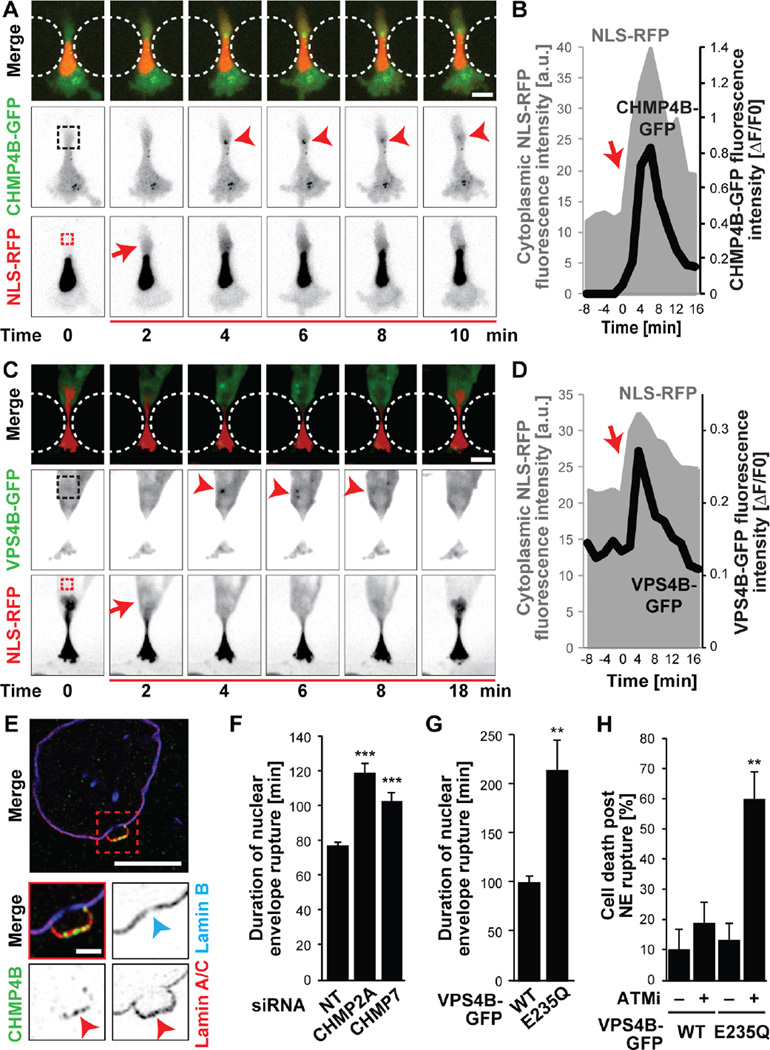
During cancer metastasis, tumor cells penetrate tissues through tight interstitial spaces, which requires extensive deformation of the cell and its nucleus. Here, we investigated mammalian tumor cell migration in confining microenvironments in vitro and in vivo. Nuclear deformation caused localized loss of nuclear envelope (NE) integrity, which led to the uncontrolled exchange of nucleo-cytoplasmic content, herniation of chromatin across the NE, and DNA damage. The incidence of NE rupture increased with cell confinement and with depletion of nuclear lamins, NE proteins that structurally support the nucleus. Cells restored NE integrity using components of the endosomal sorting complexes required for transport III (ESCRT III) machinery. Our findings indicate that cell migration incurs substantial physical stress on the NE and its content and requires efficient NE and DNA damage repair for cell survival.
Copyright © 2016, American Association for the Advancement of Science.
Figures
Comment in
-
Cell migration: Nuclear envelope ruptures as cells squeeze through tight spaces.Nat Rev Mol Cell Biol. 2016 May;17(5):263. doi: 10.1038/nrm.2016.47. Epub 2016 Apr 14. Nat Rev Mol Cell Biol. 2016. PMID: 27075412 No abstract available.
-
CELL BIOLOGY. When cells push the envelope.Science. 2016 Apr 15;352(6283):295-6. doi: 10.1126/science.aaf7735. Science. 2016. PMID: 27081057 No abstract available.
-
Cell migration: Nuclear envelope ruptures as cells squeeze through tight spaces.Nat Rev Cancer. 2016 Apr 26;16(5):272-3. doi: 10.1038/nrc.2016.42. Nat Rev Cancer. 2016. PMID: 27112202 No abstract available.
-
ESCRT machinery: Damage control at the nuclear membrane.Cell Res. 2016 Jun;26(6):641-2. doi: 10.1038/cr.2016.52. Epub 2016 May 6. Cell Res. 2016. PMID: 27151367 Free PMC article.
References
-
- Burke B, Stewart CL. Functional architecture of the cell's nucleus in development, aging, and disease. Current topics in developmental biology. 2014;109:1. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
