

Implications of Spatiotemporal Regulation of Shigella flexneri Type Three Secretion Activity on Effector Functions: Think Globally, Act Locally
- PMID: 27014638
- PMCID: PMC4783576
- DOI: 10.3389/fcimb.2016.00028
Implications of Spatiotemporal Regulation of Shigella flexneri Type Three Secretion Activity on Effector Functions: Think Globally, Act Locally
Abstract
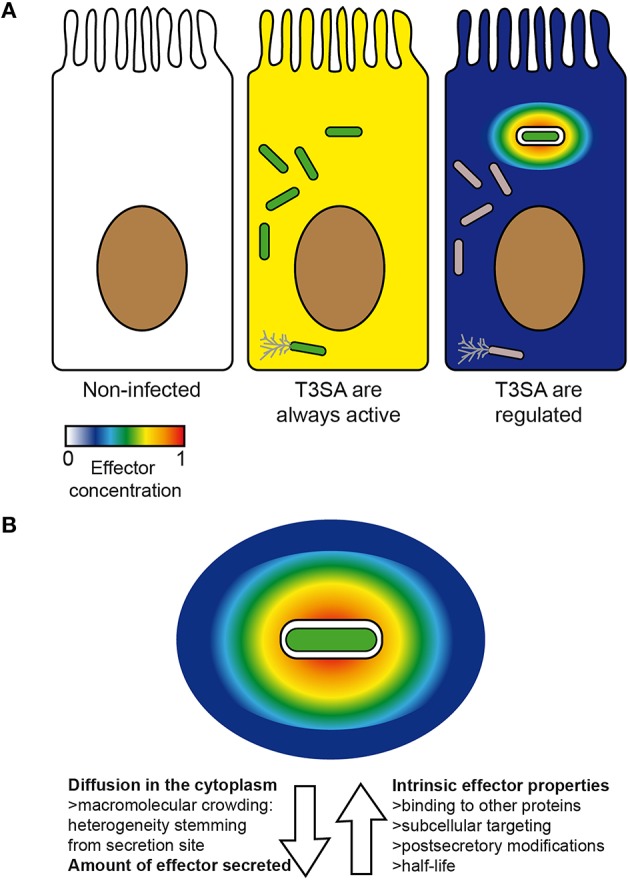
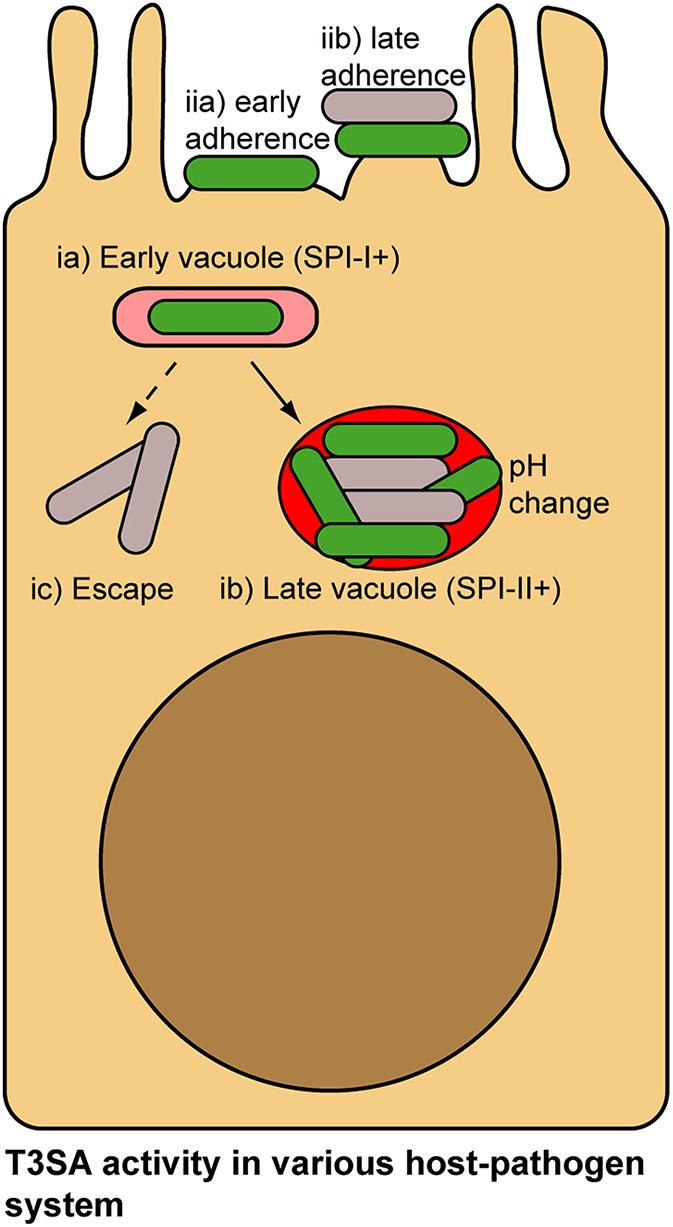
Shigella spp. are Gram-negative bacterial pathogens that infect human colonic epithelia and cause bacterial dysentery. These bacteria express multiple copies of a syringe-like protein complex, the Type Three Secretion apparatus (T3SA), which is instrumental in the etiology of the disease. The T3SA triggers the plasma membrane (PM) engulfment of the bacteria by host cells during the initial entry process. It then enables bacteria to escape the resulting phagocytic-like vacuole. Freed bacteria form actin comets to move in the cytoplasm, which provokes bacterial collision with the inner leaflet of the PM. This phenomenon culminates in T3SA-dependent secondary uptake and vacuolar rupture in neighboring cells in a process akin to what is observed during entry and named cell-to-cell spread. The activity of the T3SA of Shigella flexneri was recently demonstrated to display an on/off regulation during the infection. While the T3SA is active when bacteria are in contact with PM-derived compartments, it switches to an inactive state when bacteria are released within the cytosol. These observations indicate that effector proteins transiting through the T3SA are therefore translocated in a highly time and space constrained fashion, likely impacting on their cellular distribution. Herein, we present what is currently known about the composition, the assembly and the regulation of the T3SA activity and discuss the consequences of the on/off regulation of T3SA on Shigella effector properties and functions during the infection. Specific examples that will be developed include the role of effectors IcsB and VirA in the escape from LC3/ATG8-positive vacuoles formed during cell-to-cell spread and of IpaJ protease activity against N-miristoylated proteins. The conservation of a similar regulation of T3SA activity in other pathogens such as Salmonella or Enteropathogenic Escherichia coli will also be briefly discussed.
Keywords: Shigella; effectors; enteropathogens; host-pathogen interactions; signal transduction; type three secretion apparatus; type three secretion system.
Figures



References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous