

Developmental Cycle and Genome Analysis of "Rubidus massiliensis," a New Vermamoeba vermiformis Pathogen
- PMID: 27014641
- PMCID: PMC4791399
- DOI: 10.3389/fcimb.2016.00031
Developmental Cycle and Genome Analysis of "Rubidus massiliensis," a New Vermamoeba vermiformis Pathogen
Abstract
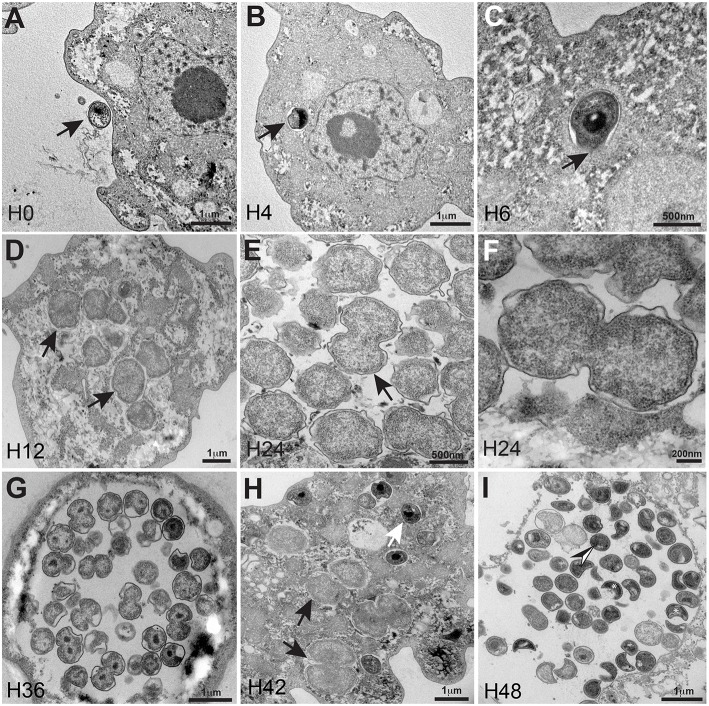
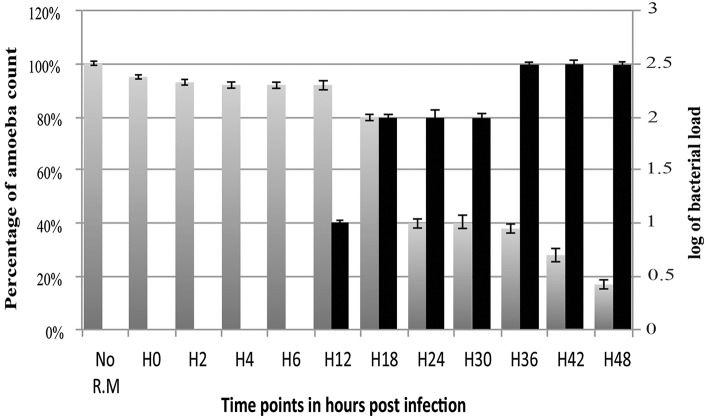
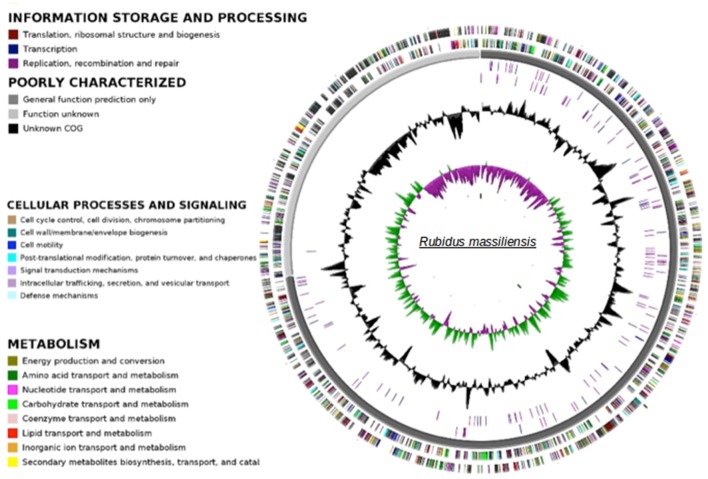
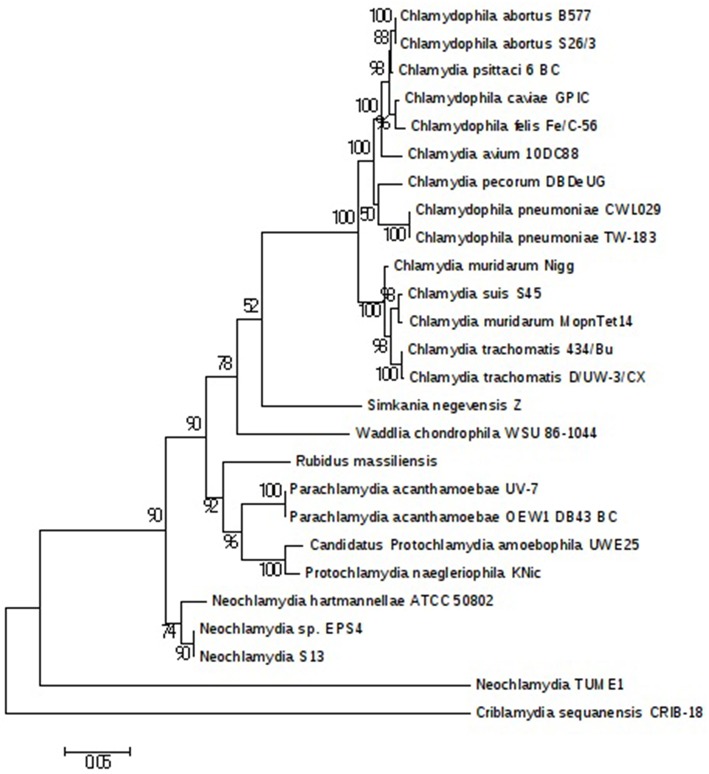
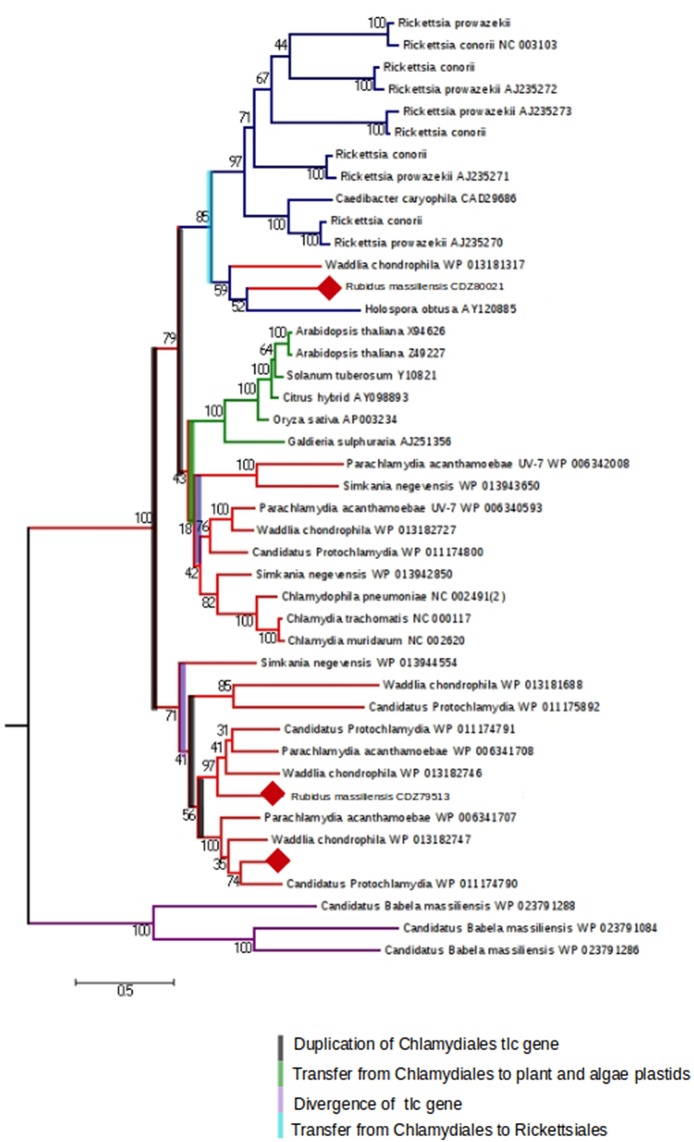
The study of amoeba-associated Chlamydiae is a dynamic field in which new species are increasingly reported. In the present work, we characterized the developmental cycle and analyzed the genome of a new member of this group associated with Vermamoeba vermiformis, we propose to name "Rubidus massiliensis." This bacterium is well-adapted to its amoeba host and do not reside inside of inclusion vacuoles after phagocytosis. It has a developmental cycle typical of this family of bacteria, with a transition from condensed elementary bodies to hypodense replicative reticulate bodies. Multiplication occurs through binary fission of the reticulate bodies. The genome of "R. massiliensis" consists of a 2.8 Mbp chromosome and two plasmids (pRm1, pRm2) consisting of 39,075 bp and 80,897 bp, respectively, a feature that is unique within this group. The Re-analysis of the Chlamydiales genomes including the one of "R. massiliensis" slightly modified the previous phylogeny of the tlc gene encoding the ADP/ATP translocase. Our analysis suggested that the tlc gene could have been transferred to plant and algal plastids before the transfer to Rickettsiales, and that this gene was probably duplicated several times.
Keywords: Rubidus massiliensis; Vermamoeba vermiformis; chlamydiae; co-culture; host specificity.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous