

Dietary Plant Lectins Appear to Be Transported from the Gut to Gain Access to and Alter Dopaminergic Neurons of Caenorhabditis elegans, a Potential Etiology of Parkinson's Disease
- PMID: 27014695
- PMCID: PMC4780318
- DOI: 10.3389/fnut.2016.00007
Dietary Plant Lectins Appear to Be Transported from the Gut to Gain Access to and Alter Dopaminergic Neurons of Caenorhabditis elegans, a Potential Etiology of Parkinson's Disease
Abstract
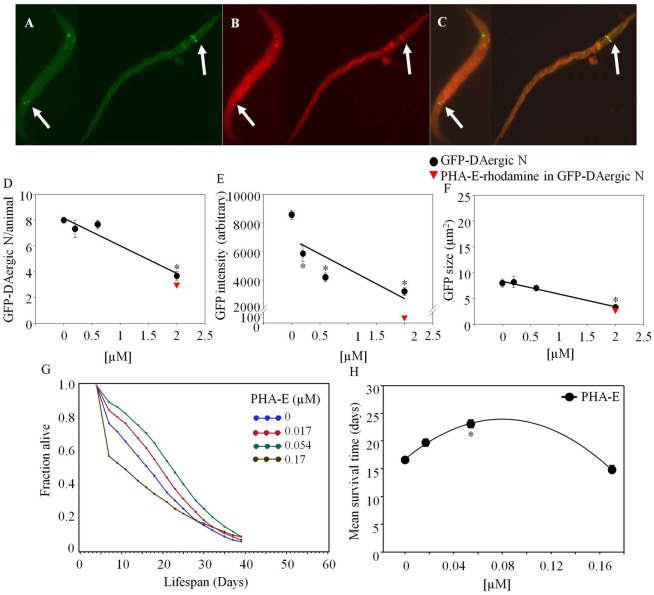
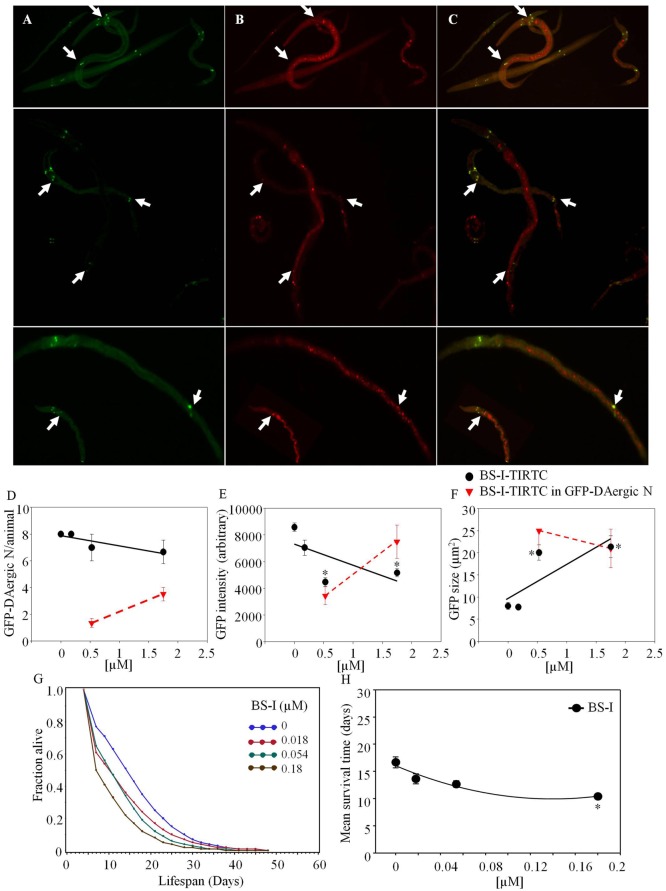
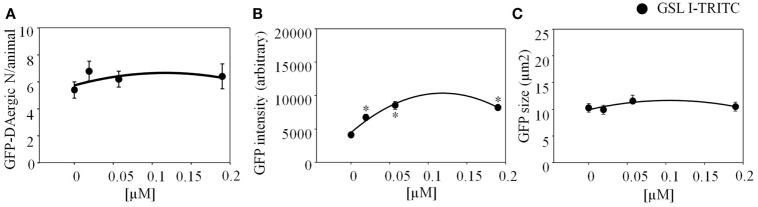
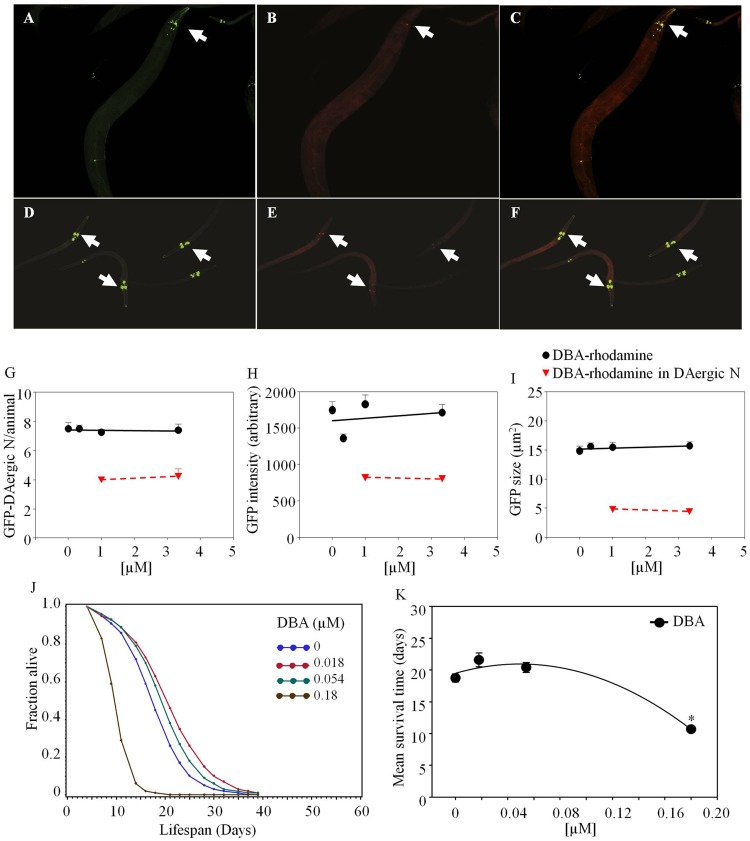
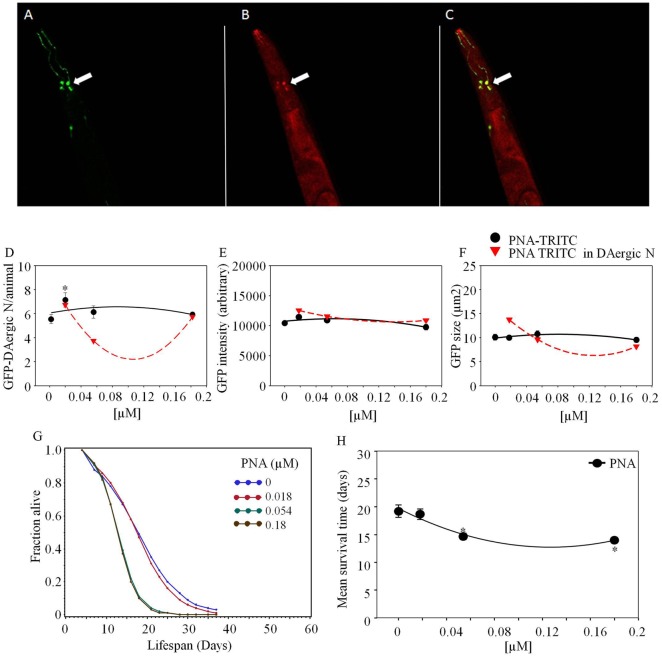
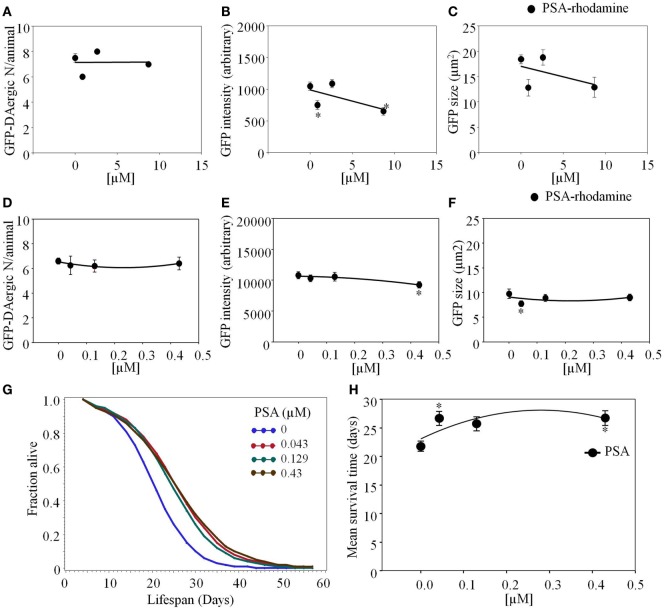
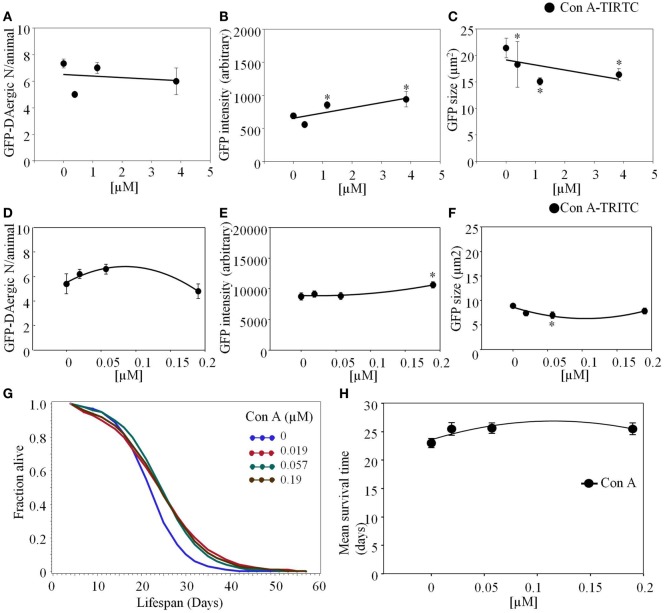
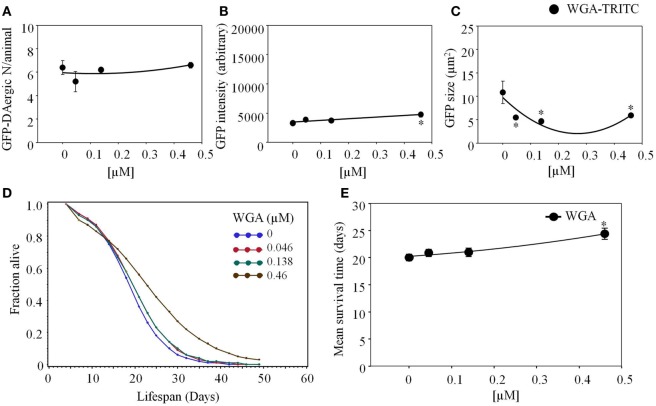
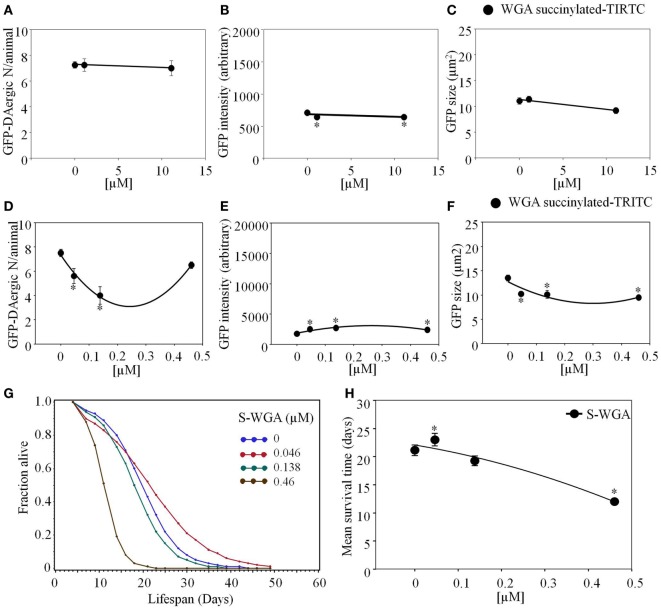
Lectins from dietary plants have been shown to enhance drug absorption in the gastrointestinal tract of rats, be transported trans-synaptically as shown by tracing of axonal and dendritic paths, and enhance gene delivery. Other carbohydrate-binding protein toxins are known to traverse the gut intact in dogs. Post-feeding rhodamine- or TRITC-tagged dietary lectins, the lectins were tracked from gut to dopaminergic neurons (DAergic-N) in transgenic Caenorhabditis elegans (C. elegans) [egIs1(Pdat-1:GFP)] where the mutant has the green fluorescent protein (GFP) gene fused to a dopamine transport protein gene labeling DAergic-N. The lectins were supplemented along with the food organism Escherichia coli (OP50). Among nine tested rhodamine/TRITC-tagged lectins, four, including Phaseolus vulgaris erythroagglutinin (PHA-E), Bandeiraea simplicifolia (BS-I), Dolichos biflorus agglutinin (DBA), and Arachis hypogaea agglutinin (PNA), appeared to be transported from gut to the GFP-DAergic-N. Griffonia Simplicifolia and PHA-E, reduced the number of GFP-DAergic-N, suggesting a toxic activity. PHA-E, BS-I, Pisum sativum (PSA), and Triticum vulgaris agglutinin (Succinylated) reduced fluorescent intensity of GFP-DAergic-N. PHA-E, PSA, Concanavalin A, and Triticum vulgaris agglutinin decreased the size of GFP-DAergic-N, while BS-I increased neuron size. These observations suggest that dietary plant lectins are transported to and affect DAergic-N in C. elegans, which support Braak and Hawkes' hypothesis, suggesting one alternate potential dietary etiology of Parkinson's disease (PD). A recent Danish study showed that vagotomy resulted in 40% lower incidence of PD over 20 years. Differences in inherited sugar structures of gut and neuronal cell surfaces may make some individuals more susceptible in this conceptual disease etiology model.
Keywords: Caenorhabditis elegans; dopamine transporter; dopaminergic neurons; fluorescence; plant lectins.
Figures
References
-
- Nachbar MS, Oppenheim JD. Lectins in the United States diet: a survey of lectins in commonly consumed foods and a review of the literature. Am J Clin Nutr (1980) 33:2338–45. - PubMed
-
- Bruylants M, Vennemann M. Le Jequirity. Bull Acad R Med Belg (1884) 3:147
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous