

Rhein Inhibits AlkB Repair Enzymes and Sensitizes Cells to Methylated DNA Damage
- PMID: 27015802
- PMCID: PMC4900258
- DOI: 10.1074/jbc.M115.711895
Rhein Inhibits AlkB Repair Enzymes and Sensitizes Cells to Methylated DNA Damage
Abstract
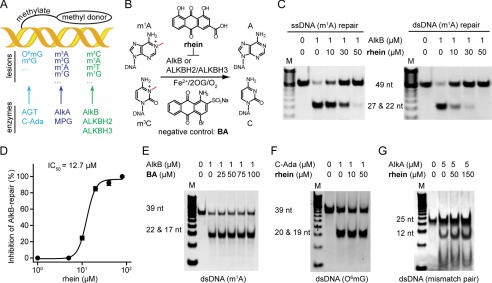
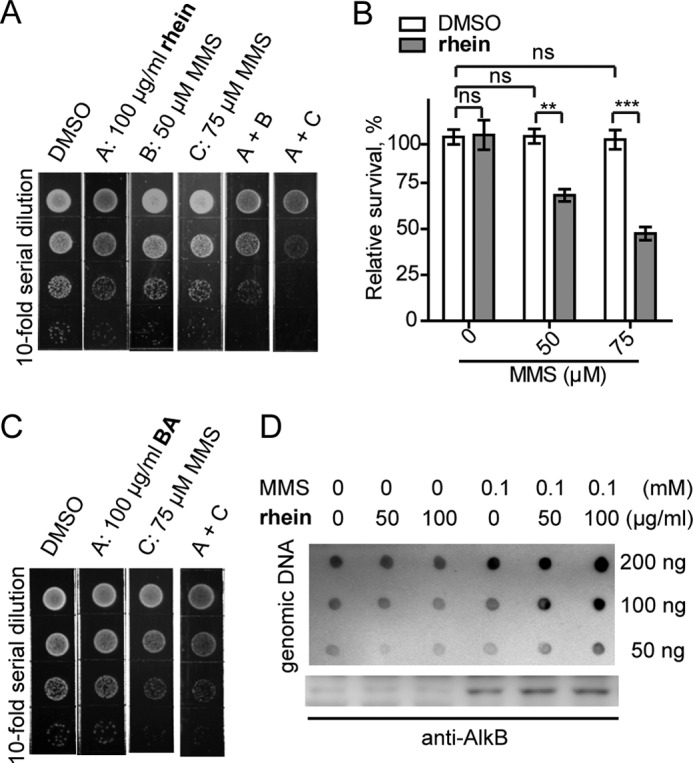
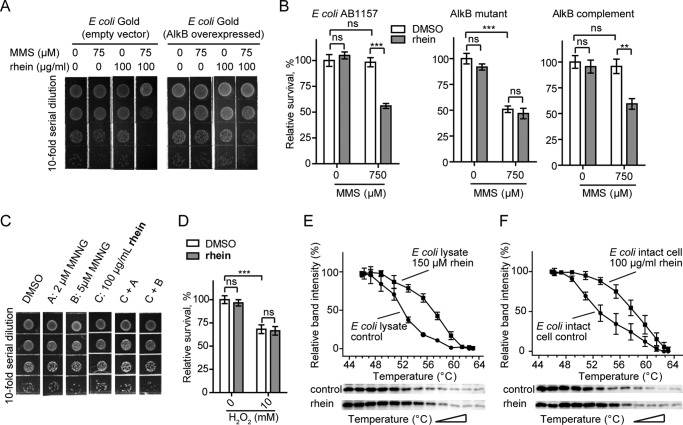
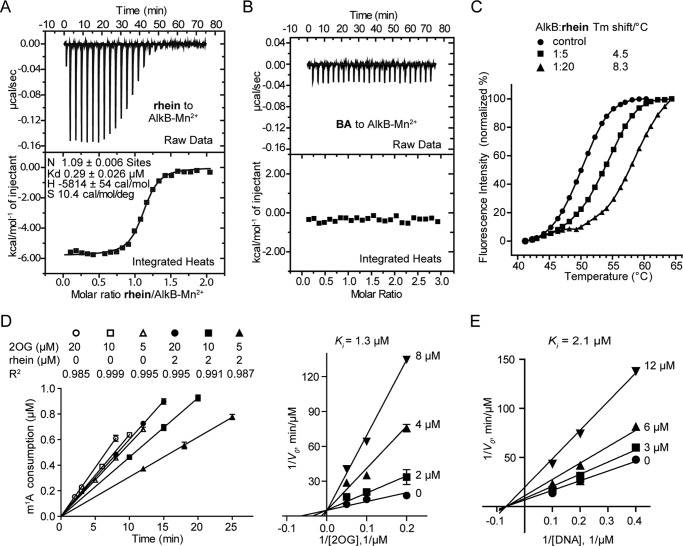
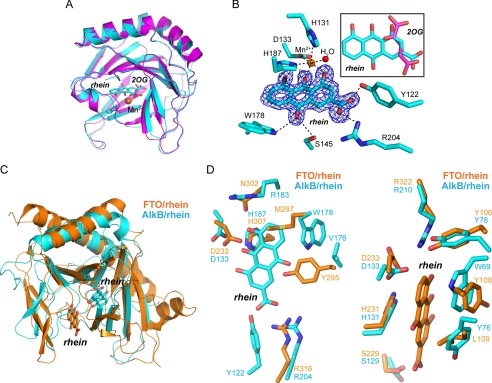
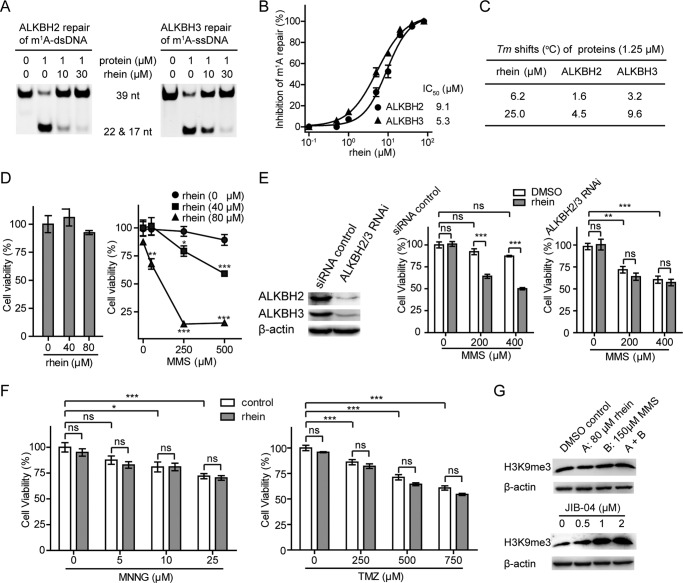
The AlkB repair enzymes, including Escherichia coli AlkB and two human homologues, ALKBH2 and ALKBH3, are iron(II)- and 2-oxoglutarate-dependent dioxygenases that efficiently repair N(1)-methyladenine and N(3)-methylcytosine methylated DNA damages. The development of small molecule inhibitors of these enzymes has seen less success. Here we have characterized a previously discovered natural product rhein and tested its ability to inhibit AlkB repair enzymes in vitro and to sensitize cells to methyl methane sulfonate that mainly produces N(1)-methyladenine and N(3)-methylcytosine lesions. Our investigation of the mechanism of rhein inhibition reveals that rhein binds to AlkB repair enzymes in vitro and promotes thermal stability in vivo In addition, we have determined a new structural complex of rhein bound to AlkB, which shows that rhein binds to a different part of the active site in AlkB than it binds to in fat mass and obesity-associated protein (FTO). With the support of these observations, we put forth the hypothesis that AlkB repair enzymes would be effective pharmacological targets for cancer treatment.
Keywords: DNA repair; DNA-protein interaction; bacteria; cancer therapy; chemical biology; enzyme inhibitor; enzyme structure.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
References
-
- Sedgwick B. (2004) Repairing DNA-methylation damage. Nat. Rev. Mol. Cell Biol. 5, 148–157 - PubMed
-
- Rose N. R., McDonough M. A., King O. N., Kawamura A., and Schofield C. J. (2011) Inhibition of 2-oxoglutarate dependent oxygenases. Chem. Soc. Rev. 40, 4364–4397 - PubMed
-
- Zhang Y., Chen F. X., Mehta P., and Gold B. (1993) Groove- and sequence-selective alkylation of DNA by sulfonate esters tethered to lexitropsins. Biochemistry 32, 7954–7965 - PubMed
-
- Shah D., and Gold B. (2003) Evidence in Escherichia coli that N3-methyladenine lesions and cytotoxicity induced by a minor groove binding methyl sulfonate ester can be modulated in vivo by netropsin. Biochemistry 42, 12610–12616 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
