

Genome-wide analysis of dorsal and ventral transcriptomes of the Xenopus laevis gastrula
- PMID: 27016259
- PMCID: PMC5033668
- DOI: 10.1016/j.ydbio.2016.02.032
Genome-wide analysis of dorsal and ventral transcriptomes of the Xenopus laevis gastrula
Abstract
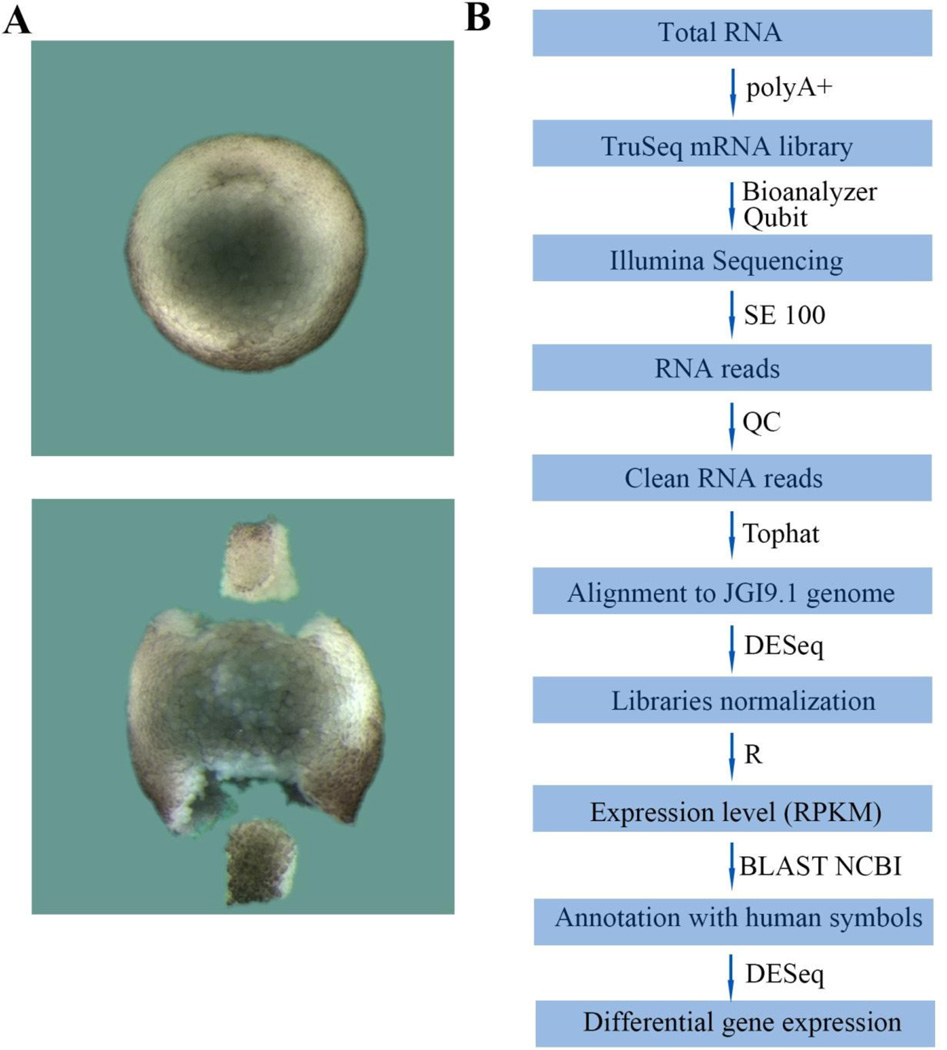
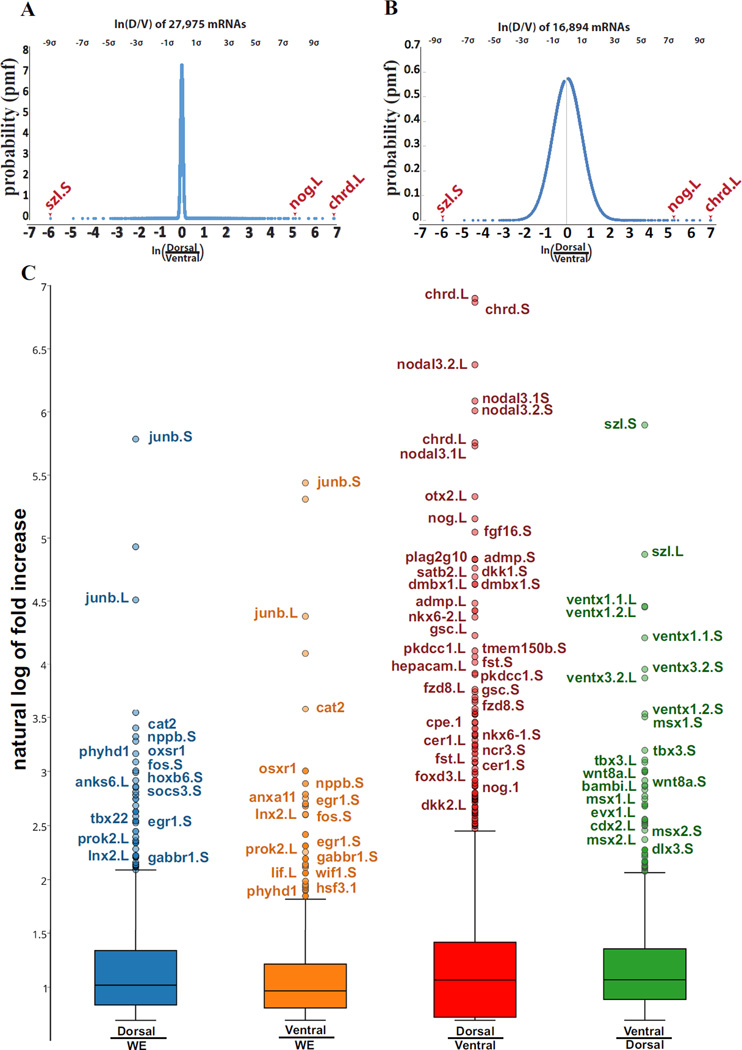
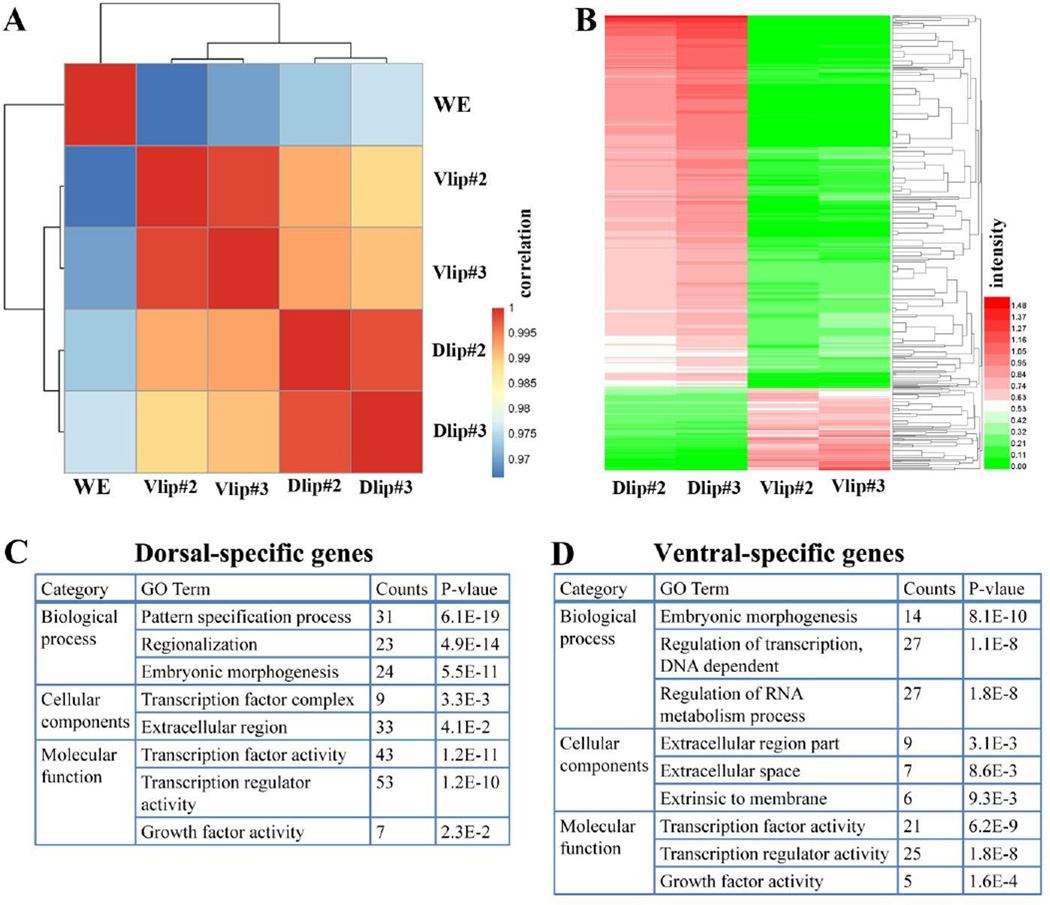
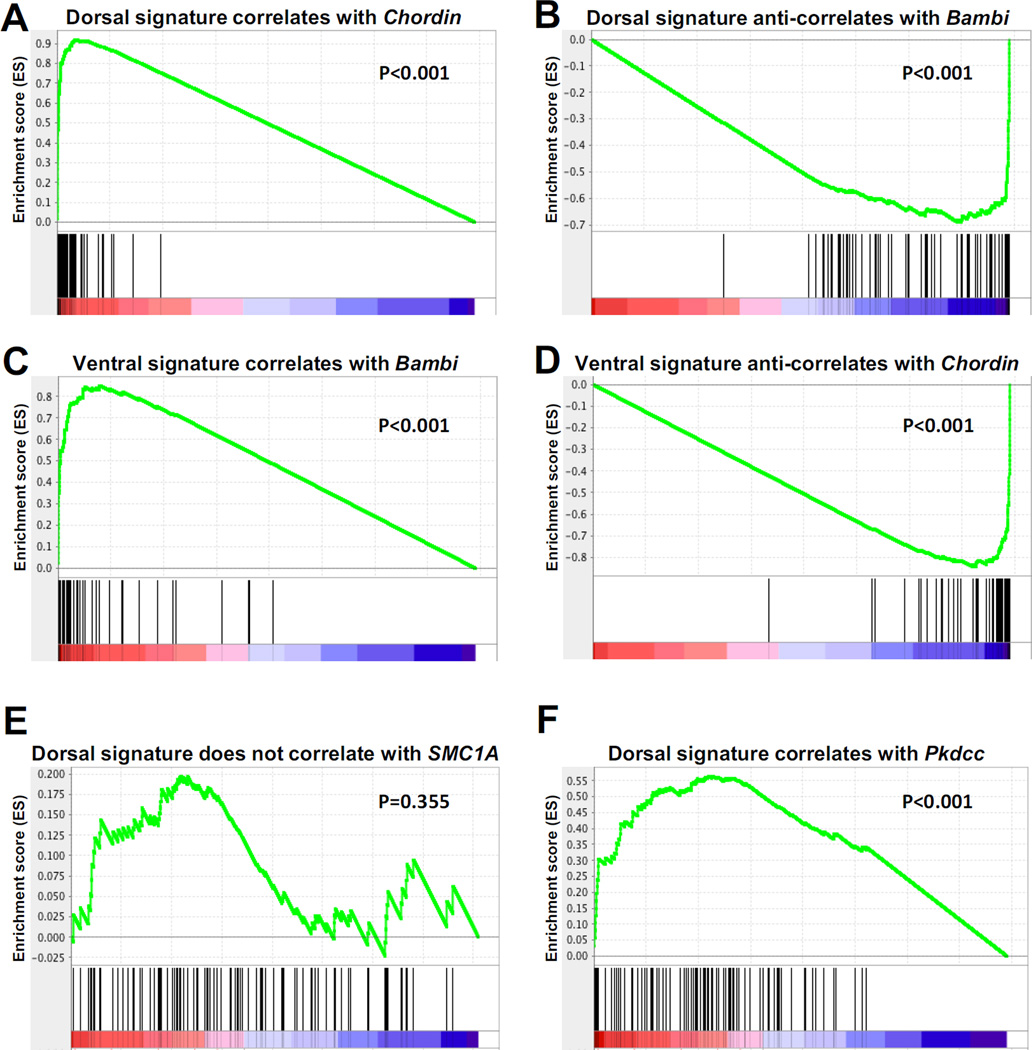
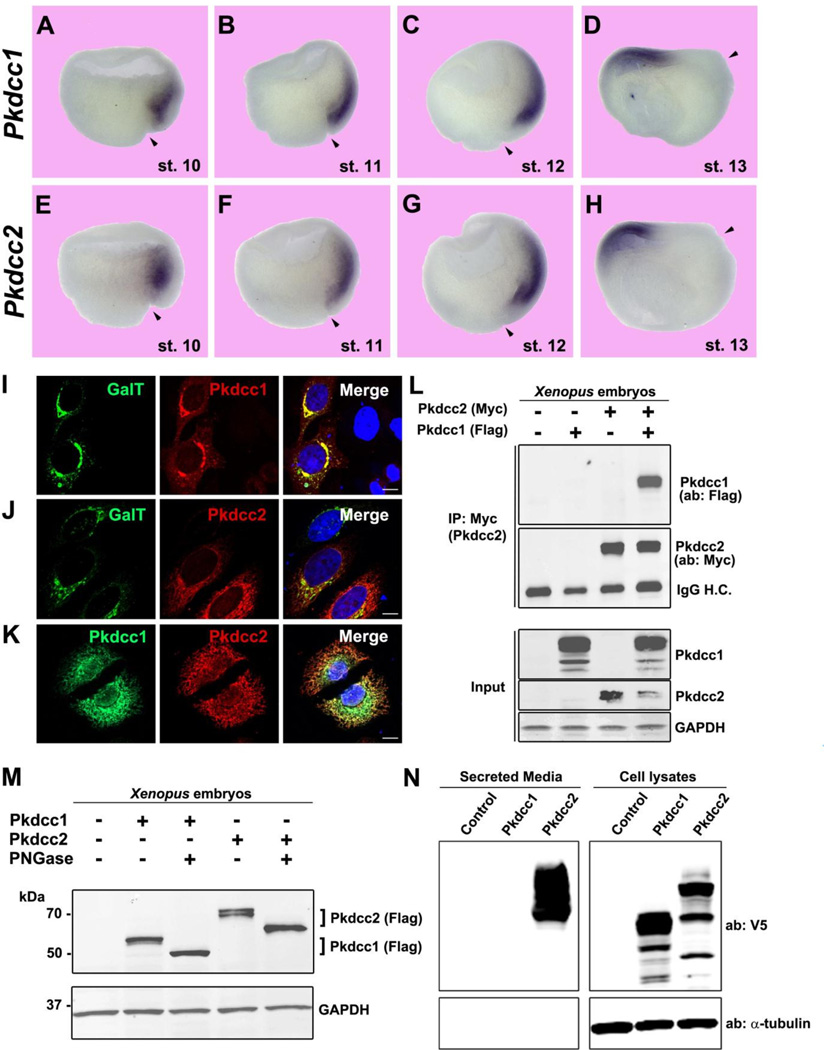
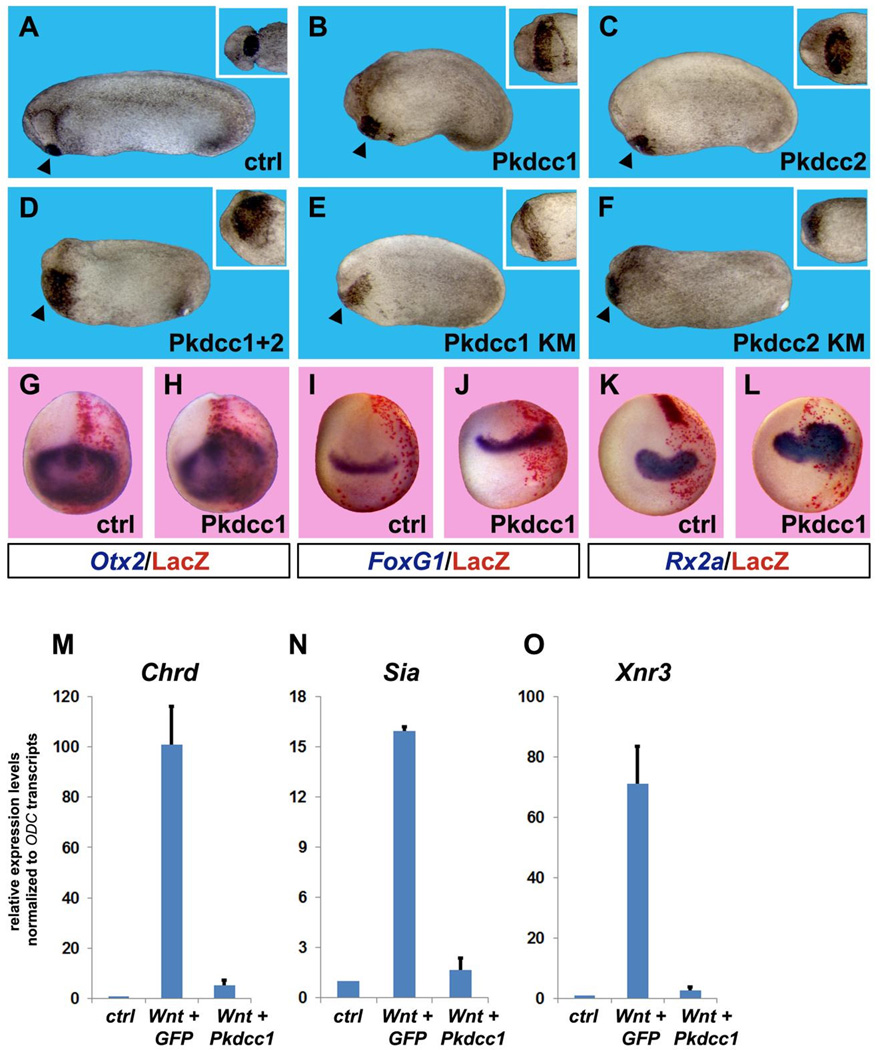
RNA sequencing has allowed high-throughput screening of differential gene expression in many tissues and organisms. Xenopus laevis is a classical embryological and cell-free extract model system, but its genomic sequence had been lacking due to difficulties arising from allotetraploidy. There is currently much excitement surrounding the release of the completed X. laevis genome (version 9.1) by the Joint Genome Institute (JGI), which provides a platform for genome-wide studies. Here we present a deep RNA-seq dataset of transcripts expressed in dorsal and ventral lips of the early Xenopus gastrula embryo using the new genomic information, which was further annotated by blast searches against the human proteome. Overall, our findings confirm previous results from differential screenings using other methods that uncovered classical dorsal genes such as Chordin, Noggin and Cerberus, as well as ventral genes such as Sizzled, Ventx, Wnt8 and Bambi. Complete transcriptome-wide tables of mRNAs suitable for data mining are presented, which include many novel dorsal- and ventral-specific genes. RNA-seq was very quantitative and reproducible, and allowed us to define dorsal and ventral signatures useful for gene set expression analyses (GSEA). As an example of a new gene, we present here data on an organizer-specific secreted protein tyrosine kinase known as Pkdcc (protein kinase domain containing, cytoplasmic) or Vlk (vertebrate lonesome kinase). Overexpression experiments indicate that Pkdcc can act as a negative regulator of Wnt/ β-catenin signaling independently of its kinase activity. We conclude that RNA-Seq in combination with the X. laevis complete genome now available provides a powerful tool for unraveling cell-cell signaling pathways during embryonic induction.
Keywords: Dorsal–Ventral patterning; RNA-Seq; Secreted Tyrosine kinase; Spemann organizer; Vlk; Xenopus laevis.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
References
-
- Andreazzoli M, Gestri G, Angeloni D, Menna E, Barsacchi G. Role of Xrx1 in Xenopus eye and anterior brain development. Development. 1999;126:2451–2460. - PubMed
-
- Bachiller D, Klingensmith J, Kemp C, Belo JA, Anderson RM, May SR, McMahon JA, McMahon AP, Harland RM, Rossant J, De Robertis EM. The organizer factors Chordin and Noggin are required for mouse forebrain development. Nature. 2000;403:658–661. - PubMed
-
- Bier E, De Robertis EM. BMP gradients: A paradigm for morphogen-mediated developmental patterning. Science. 2015;348:aaa5838. - PubMed
-
- Blum M, De Robertis EM, Wallingford JB, Niehrs C. Morpholinos: antisense and sensibility. Dev. Cell. 2015;35:145–149. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
