

Mouse model of chromosome mosaicism reveals lineage-specific depletion of aneuploid cells and normal developmental potential
- PMID: 27021558
- PMCID: PMC4820631
- DOI: 10.1038/ncomms11165
Mouse model of chromosome mosaicism reveals lineage-specific depletion of aneuploid cells and normal developmental potential
Abstract
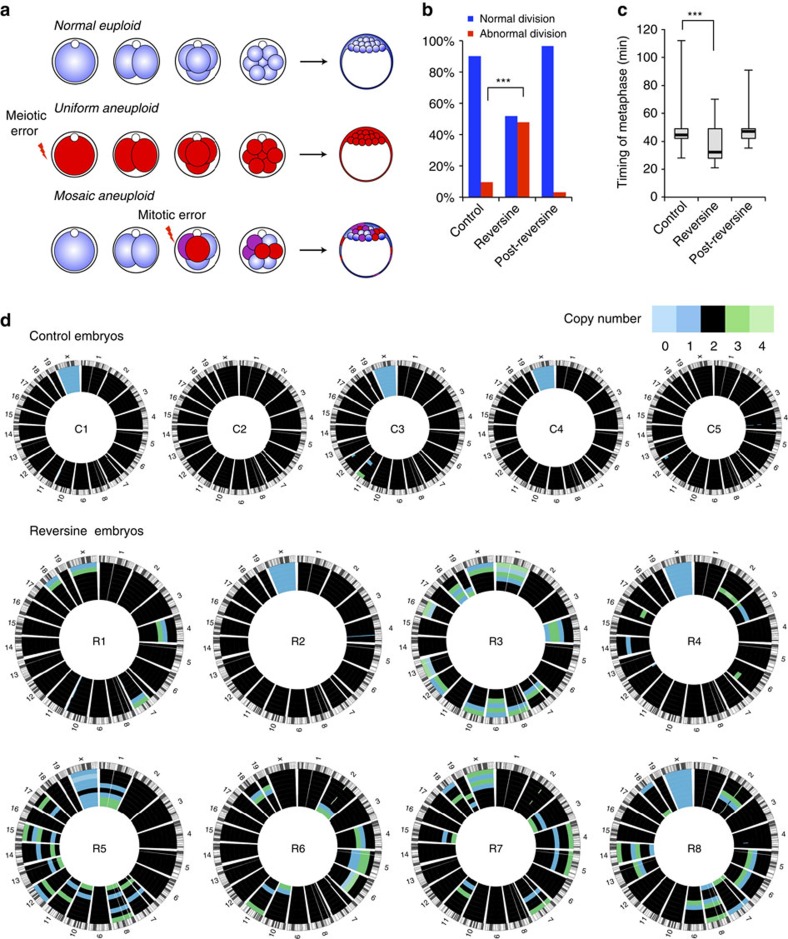
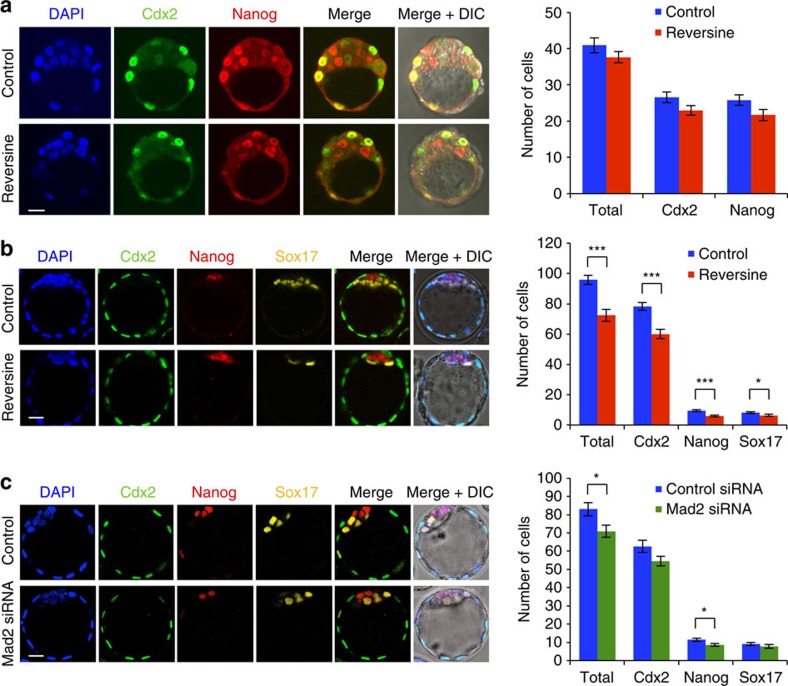
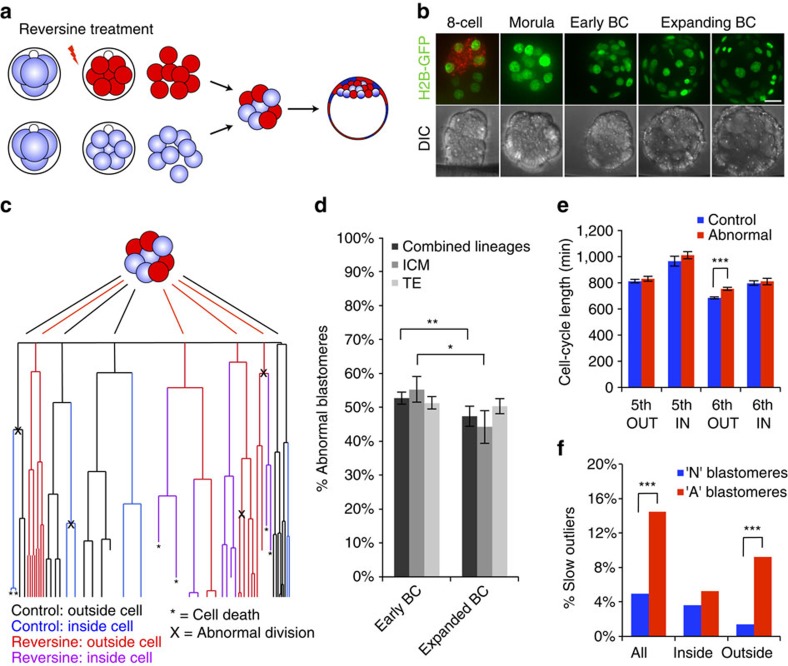
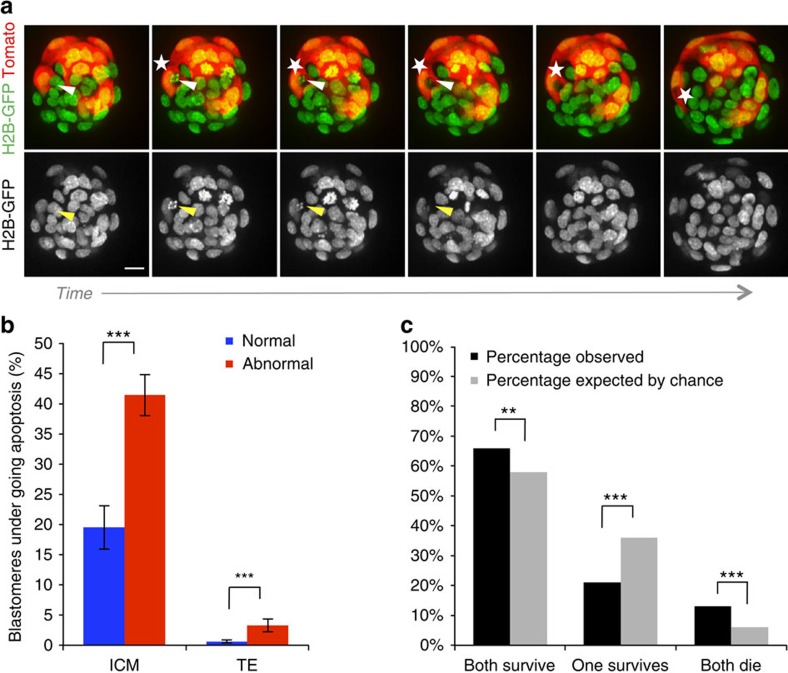
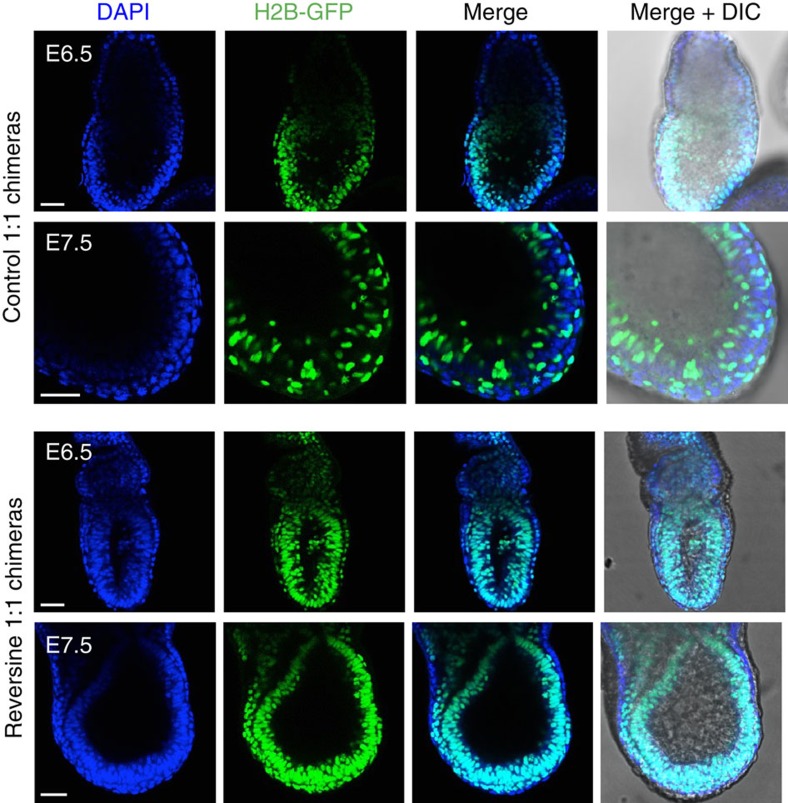
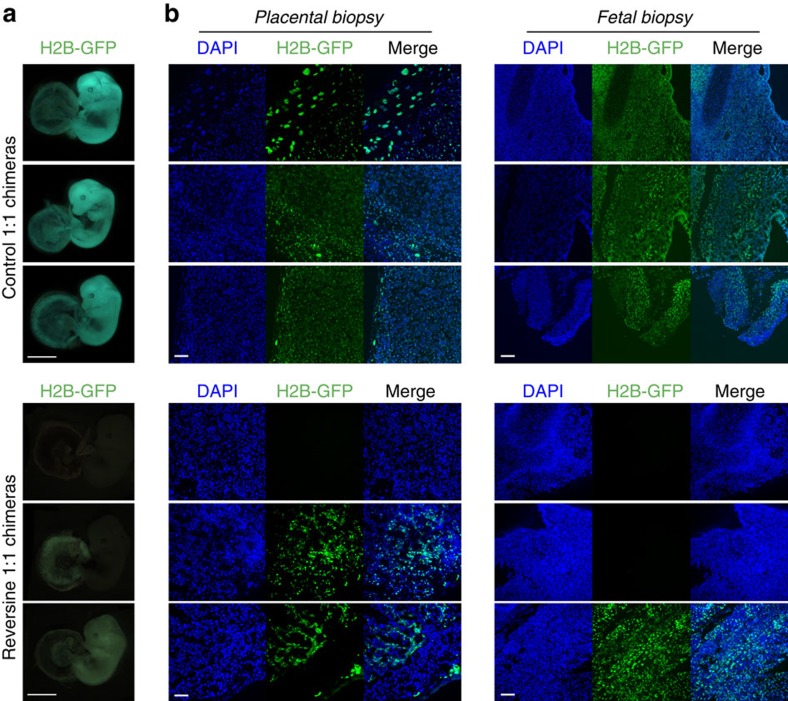
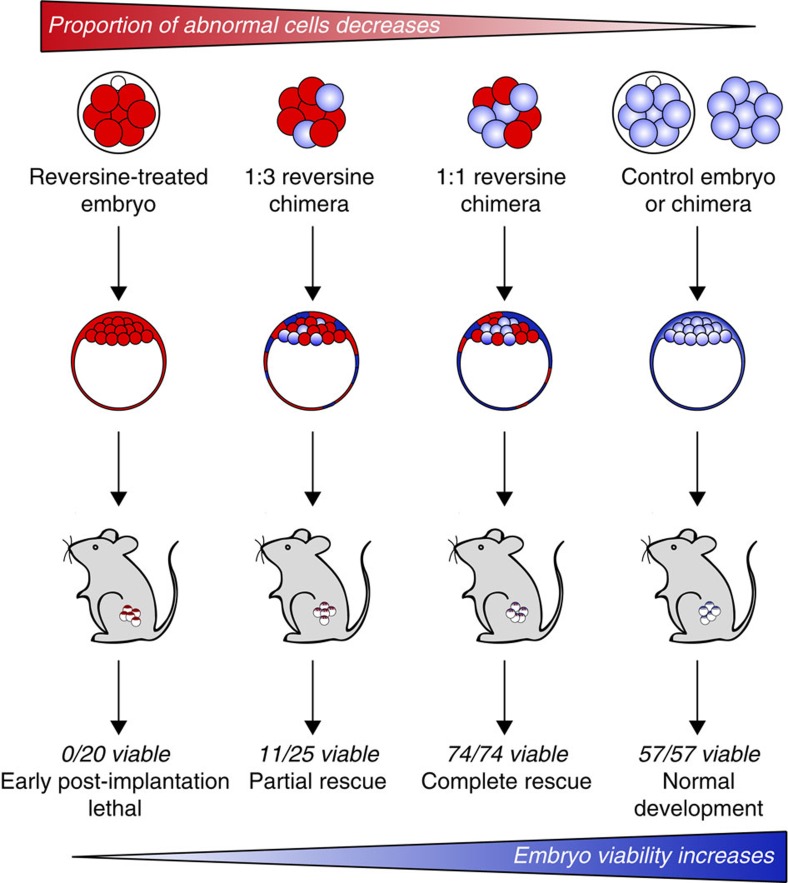
Most human pre-implantation embryos are mosaics of euploid and aneuploid cells. To determine the fate of aneuploid cells and the developmental potential of mosaic embryos, here we generate a mouse model of chromosome mosaicism. By treating embryos with a spindle assembly checkpoint inhibitor during the four- to eight-cell division, we efficiently generate aneuploid cells, resulting in embryo death during peri-implantation development. Live-embryo imaging and single-cell tracking in chimeric embryos, containing aneuploid and euploid cells, reveal that the fate of aneuploid cells depends on lineage: aneuploid cells in the fetal lineage are eliminated by apoptosis, whereas those in the placental lineage show severe proliferative defects. Overall, the proportion of aneuploid cells is progressively depleted from the blastocyst stage onwards. Finally, we show that mosaic embryos have full developmental potential, provided they contain sufficient euploid cells, a finding of significance for the assessment of embryo vitality in the clinic.
Figures
References
-
- van Echten-Arends J. et al. Chromosomal mosaicism in human preimplantation embryos: a systematic review. Hum. Reprod. Update 17, 620–627 (2011). - PubMed
-
- Macklon N. S., Geraedts J. P. M. & Fauser B. C. J. M. Conception to ongoing pregnancy: the ‘black box' of early pregnancy loss. Hum. Reprod. Update 8, 333–343 (2002). - PubMed
-
- Magli M. C. et al. Chromosome mosaicism in day 3 aneuploid embryos that develop to morphologically normal blastocysts in vitro. Hum. Reprod. 15, 1781–1786 (2000). - PubMed
-
- Mantikou E., Wong K. M., Repping S. & Mastenbroek S. Molecular origin of mitotic aneuploidies in preimplantation embryos. Biochim. Biophys. Acta 1822, 1921–1930 (2012). - PubMed
-
- Rubio C. et al. Impact of chromosomal abnormalities on preimplantation embryo development. Prenat. Diagn. 27, 748–756 (2007). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
