

The Methionine Transamination Pathway Controls Hepatic Glucose Metabolism through Regulation of the GCN5 Acetyltransferase and the PGC-1α Transcriptional Coactivator
- PMID: 27022023
- PMCID: PMC4865912
- DOI: 10.1074/jbc.M115.706200
The Methionine Transamination Pathway Controls Hepatic Glucose Metabolism through Regulation of the GCN5 Acetyltransferase and the PGC-1α Transcriptional Coactivator
Abstract
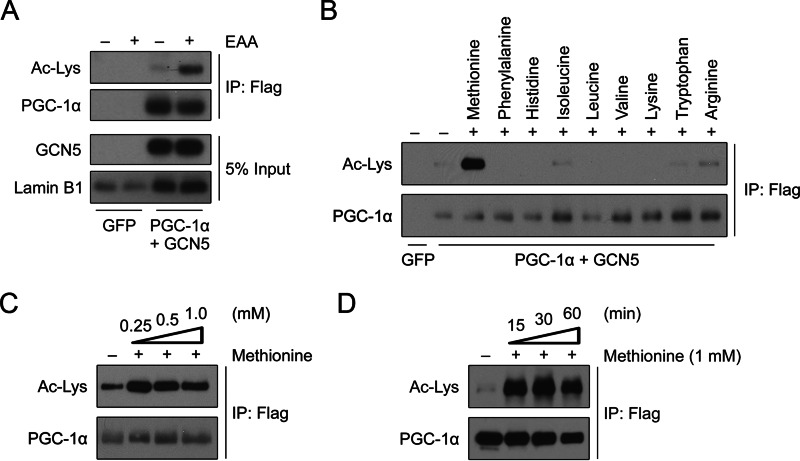
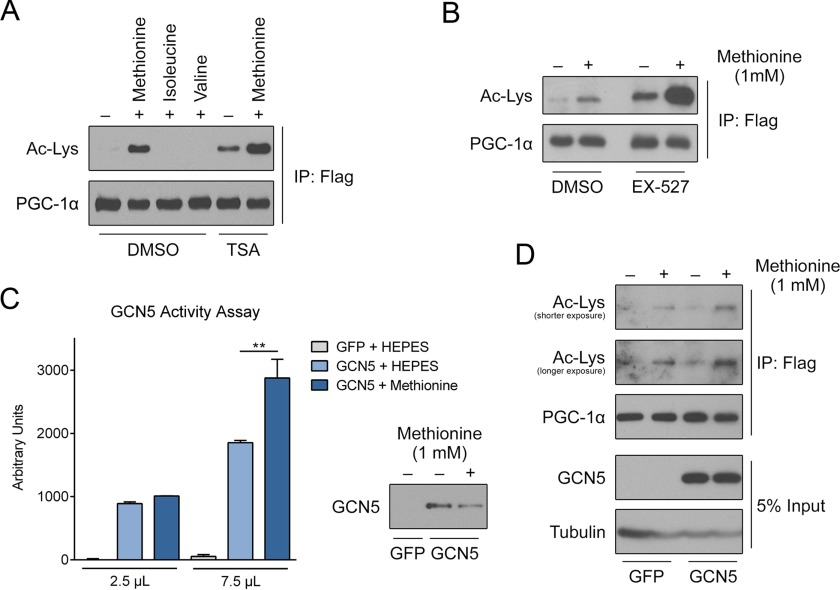
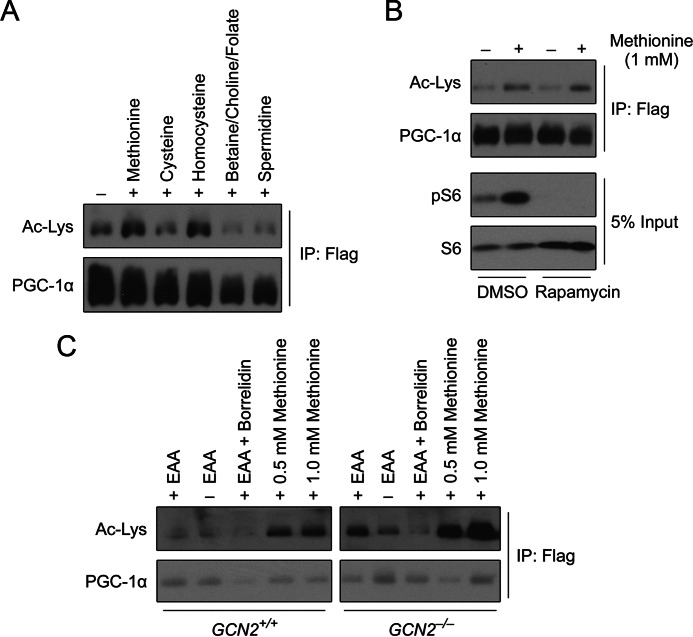
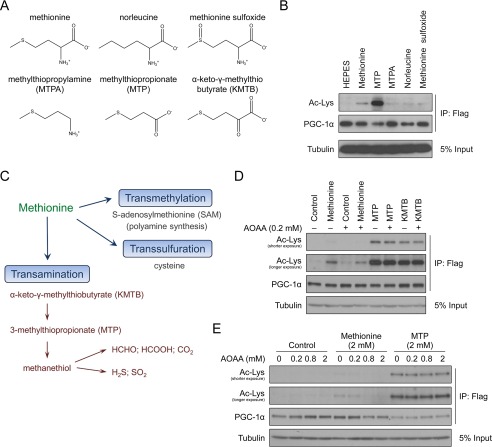
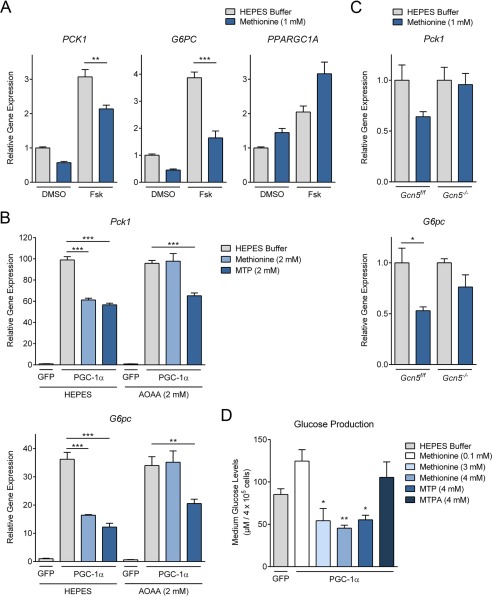
Methionine is an essential sulfur amino acid that is engaged in key cellular functions such as protein synthesis and is a precursor for critical metabolites involved in maintaining cellular homeostasis. In mammals, in response to nutrient conditions, the liver plays a significant role in regulating methionine concentrations by altering its flux through the transmethylation, transsulfuration, and transamination metabolic pathways. A comprehensive understanding of how hepatic methionine metabolism intersects with other regulatory nutrient signaling and transcriptional events is, however, lacking. Here, we show that methionine and derived-sulfur metabolites in the transamination pathway activate the GCN5 acetyltransferase promoting acetylation of the transcriptional coactivator PGC-1α to control hepatic gluconeogenesis. Methionine was the only essential amino acid that rapidly induced PGC-1α acetylation through activating the GCN5 acetyltransferase. Experiments employing metabolic pathway intermediates revealed that methionine transamination, and not the transmethylation or transsulfuration pathways, contributed to methionine-induced PGC-1α acetylation. Moreover, aminooxyacetic acid, a transaminase inhibitor, was able to potently suppress PGC-1α acetylation stimulated by methionine, which was accompanied by predicted alterations in PGC-1α-mediated gluconeogenic gene expression and glucose production in primary murine hepatocytes. Methionine administration in mice likewise induced hepatic PGC-1α acetylation, suppressed the gluconeogenic gene program, and lowered glycemia, indicating that a similar phenomenon occurs in vivo These results highlight a communication between methionine metabolism and PGC-1α-mediated hepatic gluconeogenesis, suggesting that influencing methionine metabolic flux has the potential to be therapeutically exploited for diabetes treatment.
Keywords: acetylation; acetyltransferase; gluconeogenesis; glucose-6-phosphatase (G6pc); methionine; methionine transamination; methylthiopropionic acid (MTP); peroxisome proliferator-activated receptor γ coactivator 1-α (PGC-1α) (PPARGC1α); phosphoenolpyruvate carboxykinase (Pck1); transcription coactivator.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures

Similar articles
-
CITED2 links hormonal signaling to PGC-1α acetylation in the regulation of gluconeogenesis.Nat Med. 2012 Mar 18;18(4):612-7. doi: 10.1038/nm.2691. Nat Med. 2012. PMID: 22426420
-
GCN5 acetyltransferase complex controls glucose metabolism through transcriptional repression of PGC-1alpha.Cell Metab. 2006 Jun;3(6):429-38. doi: 10.1016/j.cmet.2006.04.013. Cell Metab. 2006. PMID: 16753578
-
The deacetylase Sirt6 activates the acetyltransferase GCN5 and suppresses hepatic gluconeogenesis.Mol Cell. 2012 Dec 28;48(6):900-13. doi: 10.1016/j.molcel.2012.09.030. Epub 2012 Nov 8. Mol Cell. 2012. PMID: 23142079 Free PMC article.
-
GCN5 acetyltransferase in cellular energetic and metabolic processes.Biochim Biophys Acta Gene Regul Mech. 2021 Feb;1864(2):194626. doi: 10.1016/j.bbagrm.2020.194626. Epub 2020 Aug 19. Biochim Biophys Acta Gene Regul Mech. 2021. PMID: 32827753 Free PMC article. Review.
-
CAT in the HAT: catabolic inhibition by the histone acetyltransferase GCN5.Cell Metab. 2006 Jun;3(6):387-8. doi: 10.1016/j.cmet.2006.05.006. Cell Metab. 2006. PMID: 16753572 Review.
Cited by
-
Replacing Soybean Meal with Distillers Dried Grains with Solubles plus Rumen-Protected Lysine and Methionine: Effects on Growth Performance, Nutrients Digestion, Rumen Fermentation, and Serum Parameters in Hu Sheep.Animals (Basel). 2021 Aug 18;11(8):2428. doi: 10.3390/ani11082428. Animals (Basel). 2021. PMID: 34438885 Free PMC article.
-
Pathological role of methionine in the initiation and progression of biliary atresia.Front Pediatr. 2023 Sep 12;11:1263836. doi: 10.3389/fped.2023.1263836. eCollection 2023. Front Pediatr. 2023. PMID: 37772039 Free PMC article. Review.
-
Acetyltransferase GCN5 regulates autophagy and lysosome biogenesis by targeting TFEB.EMBO Rep. 2020 Jan 7;21(1):e48335. doi: 10.15252/embr.201948335. Epub 2019 Nov 21. EMBO Rep. 2020. PMID: 31750630 Free PMC article.
-
Metabolic Fingerprint of Turner Syndrome.J Clin Med. 2020 Mar 2;9(3):664. doi: 10.3390/jcm9030664. J Clin Med. 2020. PMID: 32131408 Free PMC article.
-
Methionine-Mediated Regulation of Intestinal Lipid Transportation Induced by High-Fat Diet in Rice Field Eel (Monopterus Albus).Aquac Nutr. 2023 Mar 16;2023:5533414. doi: 10.1155/2023/5533414. eCollection 2023. Aquac Nutr. 2023. PMID: 36967810 Free PMC article.
References
-
- Gietzen D. W., and Rogers Q. R. (2006) Nutritional homeostasis and indispensable amino acid sensing: a new solution to an old puzzle. Trends Neurosci. 29, 91–99 - PubMed
-
- Dong J., Qiu H., Garcia-Barrio M., Anderson J., and Hinnebusch A. G. (2000) Uncharged tRNA activates GCN2 by displacing the protein kinase moiety from a bipartite tRNA-binding domain. Mol. Cell 6, 269–279 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
