

Bone marrow adipose tissue: formation, function and regulation
- PMID: 27022859
- PMCID: PMC5351553
- DOI: 10.1016/j.coph.2016.03.001
Bone marrow adipose tissue: formation, function and regulation
Abstract
The human body requires an uninterrupted supply of energy to maintain metabolic homeostasis and energy balance. To sustain energy balance, excess consumed calories are stored as glycogen, triglycerides and protein, allowing the body to continue to function in states of starvation and increased energy expenditure. Adipose tissue provides the largest natural store of excess calories as triglycerides and plays an important role as an endocrine organ in energy homeostasis and beyond. This short review is intended to detail the current knowledge of the formation and role of bone marrow adipose tissue (MAT), a largely ignored adipose depot, focussing on the role of MAT as an endocrine organ and highlighting the pharmacological agents that regulate MAT.
Copyright © 2016 Elsevier Ltd. All rights reserved.
Figures

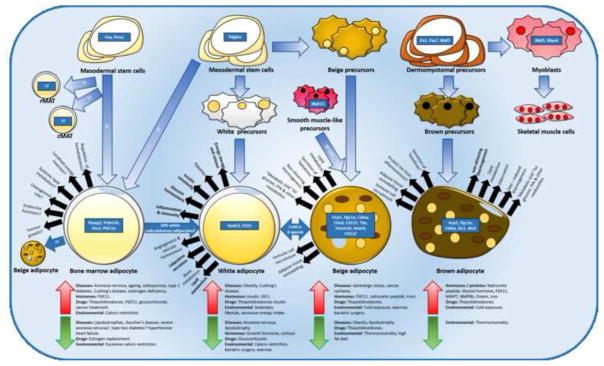
References
-
- Feller DD, Feist E. Metabolism of adipose tissue. II. Incorporation of propionate carbon into lipides by slices of adipose tissue. J Biol Chem. 1957;228:275–284. - PubMed
-
- Fruhbeck G. Overview of adipose tissue and its role in obesity and metabolic disorders. Methods Mol Biol. 2008;456:1–22. - PubMed
-
- Chau Y-Y, Bandiera R, Serrels A, Martínez-Estrada OM, Qing W, Lee M, Slight J, Thornburn A, Berry R, McHaffie S, et al. Visceral and subcutaneous fat have different origins and evidence supports a mesothelial source. Nat Cell Biol. 2014;16:367–375. The autohors demonstrate that there is a major ontogenetic difference between visceral and subcutaneous white adipose tissue. - PMC - PubMed
-
- Wajchenberg BL. Subcutaneous and visceral adipose tissue: their relation to the metabolic syndrome. Endocr Rev. 2000;21:697–738. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
