

Review
doi: 10.1146/annurev-pathol-012414-040349.
Epub 2016 Mar 17.
The Roles of MDM2 and MDMX in Cancer
Affiliations
- PMID: 27022975
- PMCID: PMC6028239
- DOI: 10.1146/annurev-pathol-012414-040349
Item in Clipboard
Review
The Roles of MDM2 and MDMX in Cancer
Annu Rev Pathol.
.
Abstract
For more than 25 years, MDM2 and its homolog MDMX (also known as MDM4) have been shown to exert oncogenic activity. These two proteins are best understood as negative regulators of the p53 tumor suppressor, although they may have additional p53-independent roles. Understanding the dysregulation of MDM2 and MDMX in human cancers and how they function either together or separately in tumorigenesis may improve methods of diagnosis and for assessing prognosis. Targeting the proteins themselves, or their regulators, may be a promising therapeutic approach to treating some forms of cancer.
Keywords: MDM2; MDMX; cancer; p53; tumorigenesis.
Figures
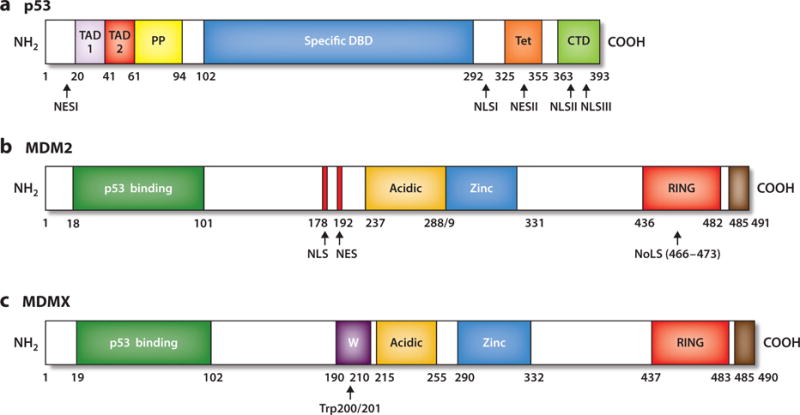
Landmarks of p53, MDM2, and MDMX proteins. (a) The N-terminal region of p53 (residues 1–101) contains two distinct transactivation domains, TAD 1 and TAD 2; a proline-rich region (PP) with five PXXP sequence motifs that are essential for its ability to induce apoptosis; and a nuclear export signal (NES) between residues 11 and 27. The central core domain (amino acids 102–292) spans the sequence-specific DNA-binding domain (DBD). The majority of missense mutations in the p53 gene are located within this domain with highly conserved subregions. The carboxyl terminus of p53 (residues 292–393) contains a flexible linker region (residues 292–324) that connects the core domain to the tetramerization domain (Tet, residues 325–355) and a basic regulatory domain (CTD, residues 363–393). The carboxyl terminus also encompasses both NES and nuclear localization signal (NLS) sequences. (b) MDM2 landmarks include the N-terminal p53-binding domain (residues 18–101); the centrally located acidic domain that binds ARF and also has a role in the ubiquitination of p53 (residues 237–288); and the zinc-finger motif, where ribosomal proteins interact (residues 289–330). The MDM2 carboxyl terminus contains a RING domain that displays E3 ubiquitin ligase activity (residues 436–482), and the carboxyl terminus tail (residues 485–491) regulates the RING motif via MDM2 homodimer formation as well as MDM2 and MDMX heterodimerization. MDM2 has NLS, nucleolar localization signal (NoLS), and NES sequences. (c) MDMX has structural domain similarity to MDM2, with an N-terminal p53 binding region (residues 19–102), central acidic (residues 215–255) and zinc-finger (residues 290–331) regions, and a carboxyl-terminally located RING domain (residues 437–483). MDMX lacks NLS, NES, or NoLS sequences but has a unique WWW motif (W, residues 190–210), which inhibits interaction with p53. In contrast to the MDM2 RING E3 ligase domain, the MDMX RING domain lacks E3 ligase activity. The carboxyl-terminal tail (residues 485–490) of MDMX is required for formation of MDM2–MDMX heterodimers.
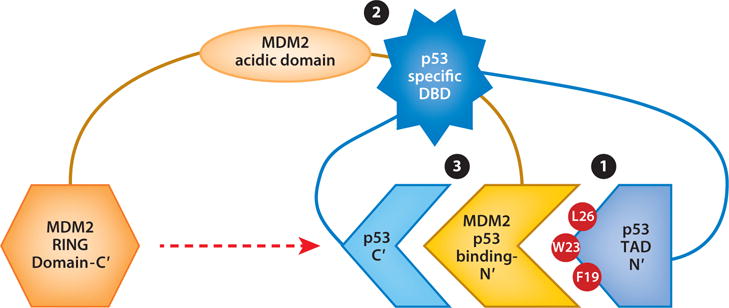
MDM2–p53 interactions involve three regions of each protein. (❶) The primary interaction regions are within the MDM2 N-terminal domain and the p53 N-terminal transactivation domain, TAD 1. (❷) The central regions of MDM2 (acidic domain) interact with the sequence-specific DNA-binding domain (DBD) of p53. (❸) The MDM2 N-terminal domain interacts with the p53 carboxyl-terminal domain, which harbors lysines whose ubiquitination requires the MDM2 RING domain (red dashed arrow). Abbreviations: C′, carboxyl terminus; F, phenylalanine; L, leucine; N′, N terminus; W, tryptophan.
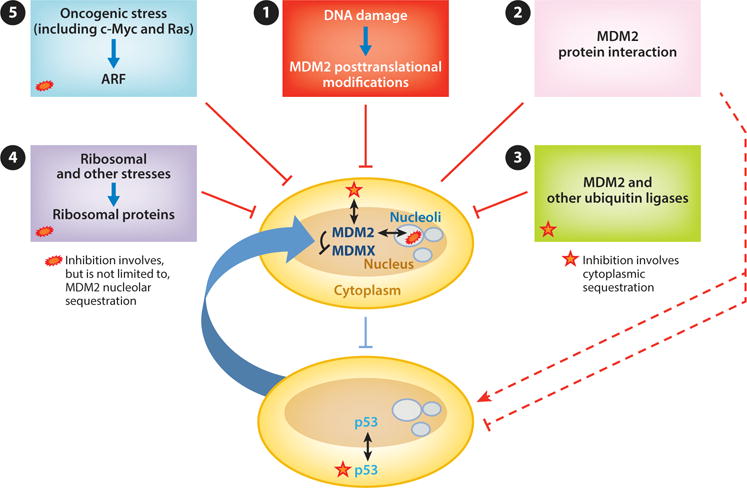
Regulation of the MDM2– and MDMX–p53 axis. In clockwise order from top: (❶) DNA damage signaling leads to posttranslational modifications of MDM2 that inhibit its interaction with and degradation of p53. Some forms of DNA damage also lead to MDM2 degradation of MDMX, thereby further activating p53. (❷) Numerous proteins interact with MDM2 and regulate its E3 ligase ability to activate or repress p53 (red dashed lines). (❸) MDM2 is degraded by as yet unidentified E3 ligases as well as by MDM2 itself in some settings. (❹) Ribosomal and other stresses cause the release of ribosomal proteins that bind and inhibit MDM2. (❺) Oncogenic stressors (including c-Myc and Ras) cause induction of ARF, which binds MDM2 and can either inhibit its E3 ligase activity or facilitate its translocation to the nucleolus.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
