

Essential role of the TFIID subunit TAF4 in murine embryogenesis and embryonic stem cell differentiation
- PMID: 27026076
- PMCID: PMC4820908
- DOI: 10.1038/ncomms11063
Essential role of the TFIID subunit TAF4 in murine embryogenesis and embryonic stem cell differentiation
Abstract
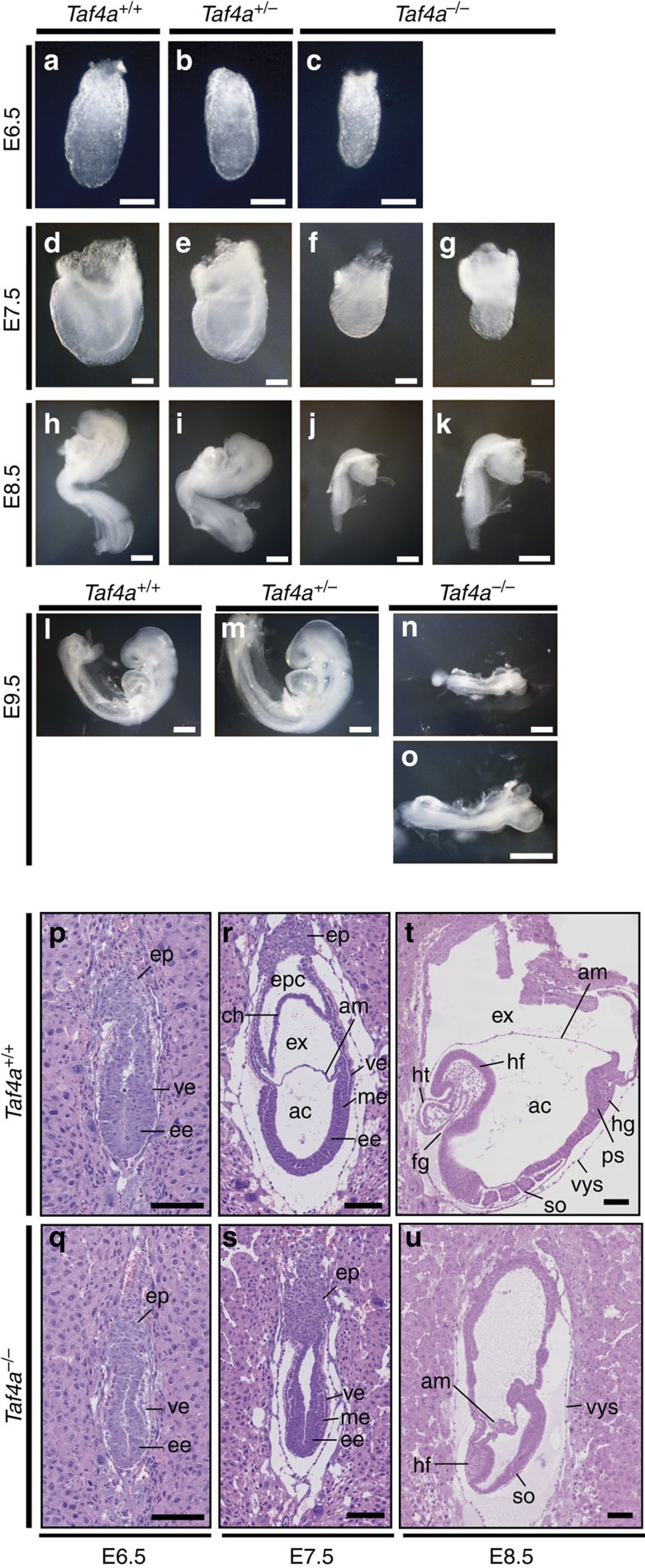
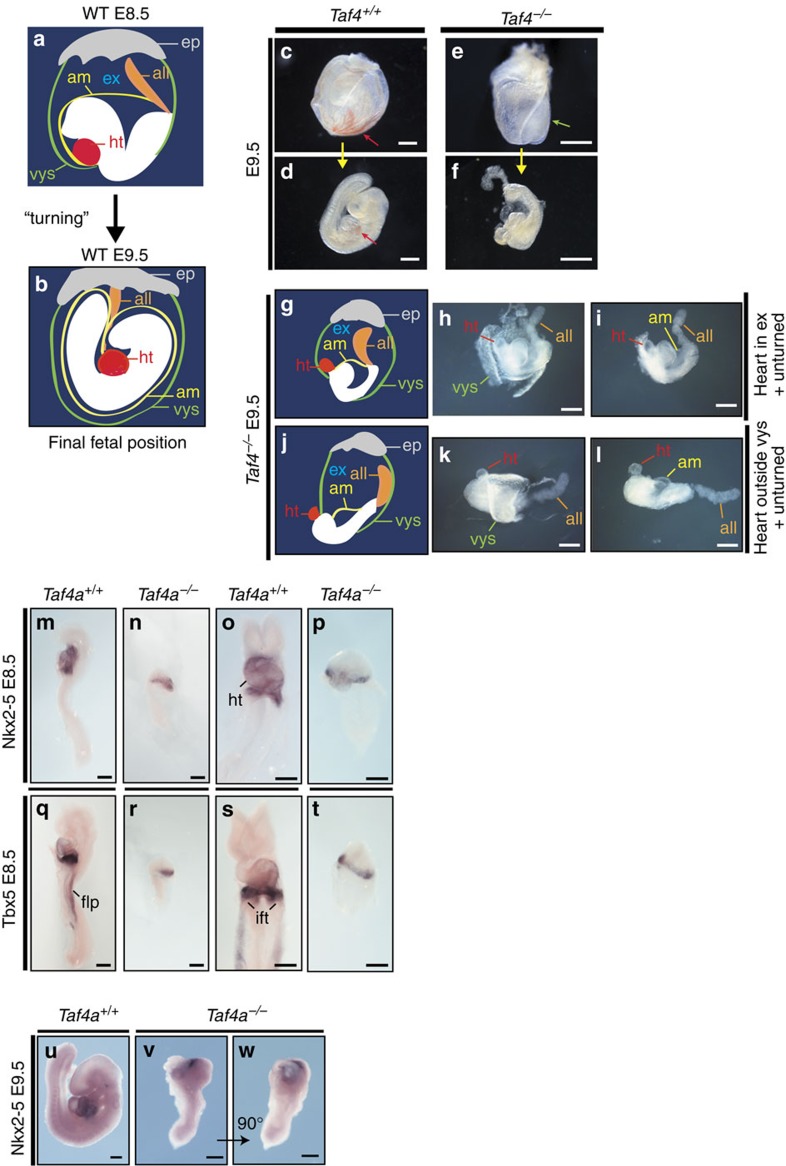
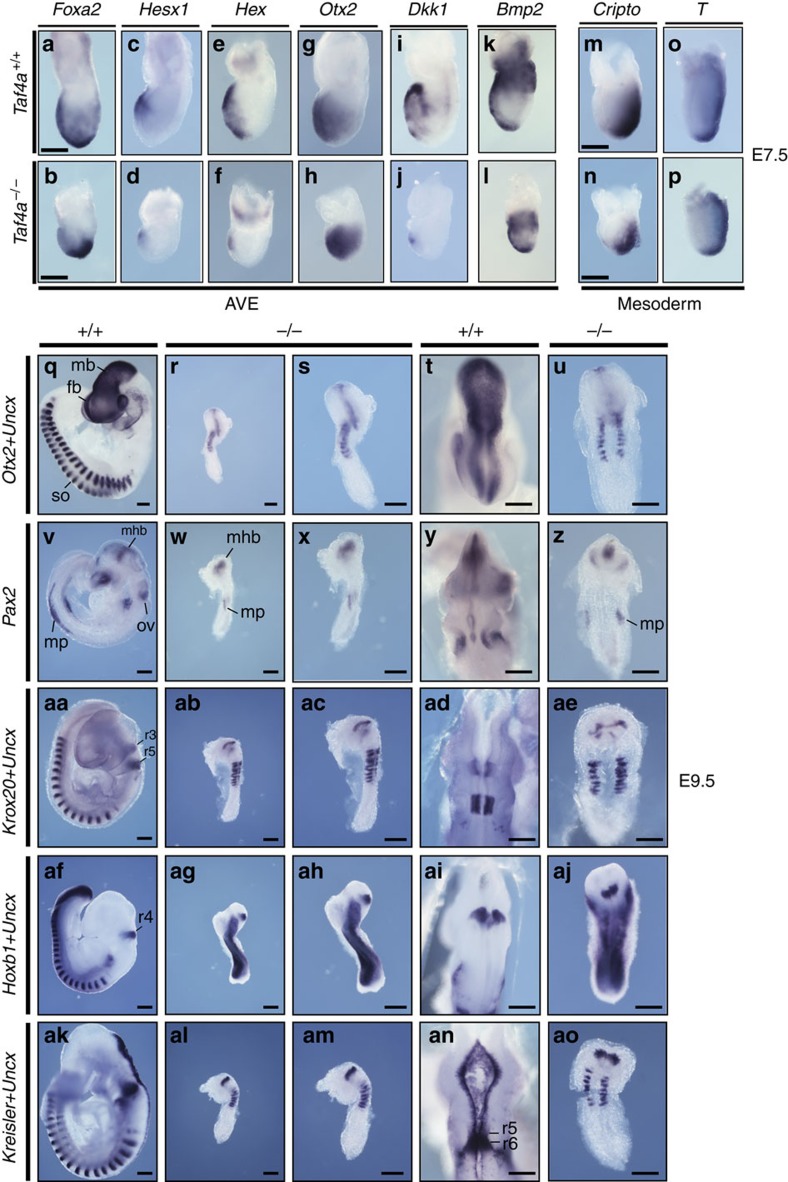
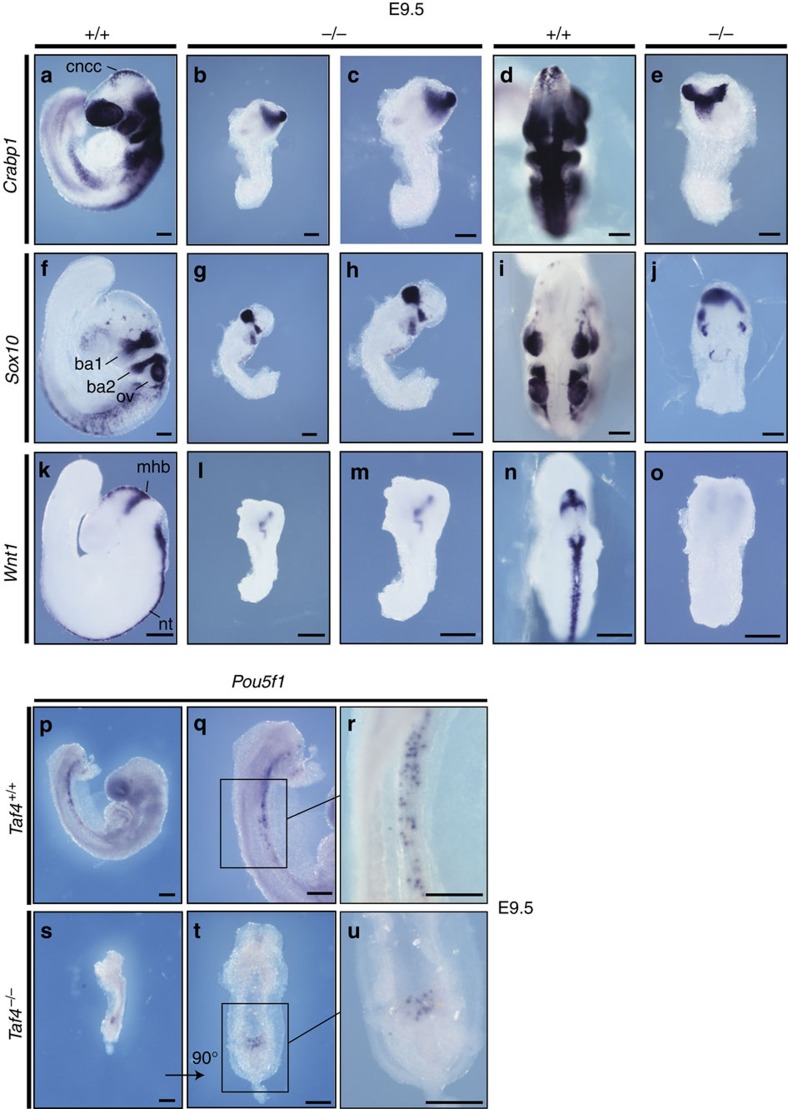
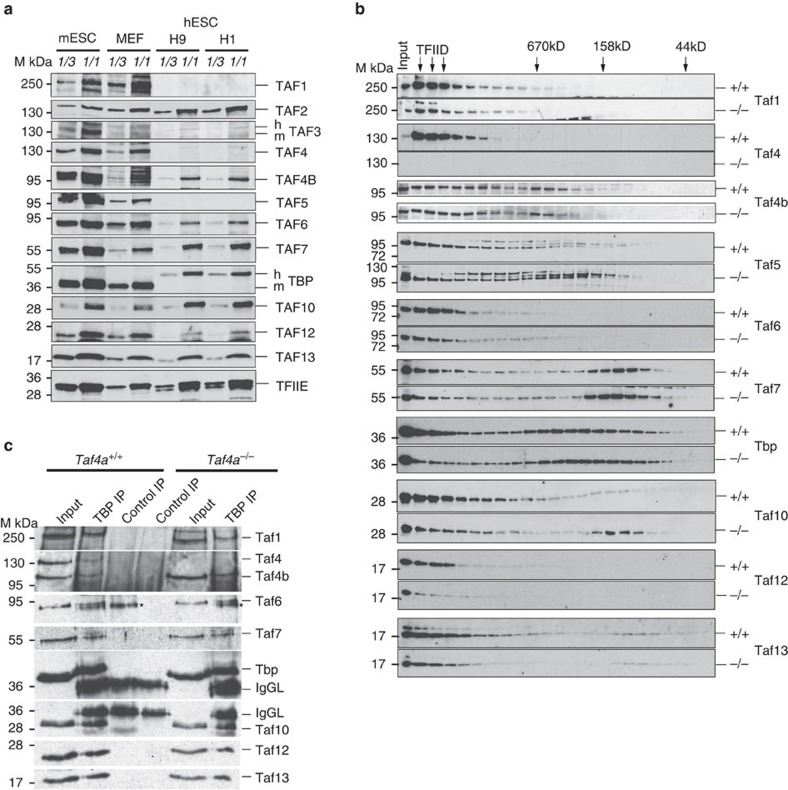
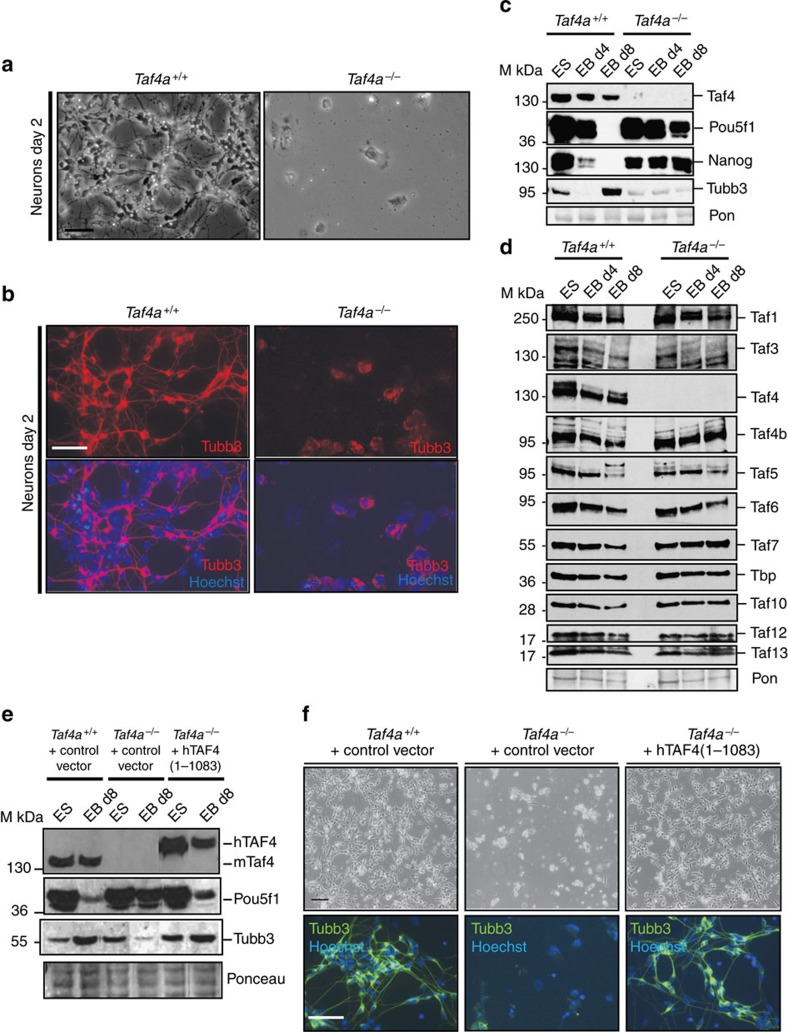
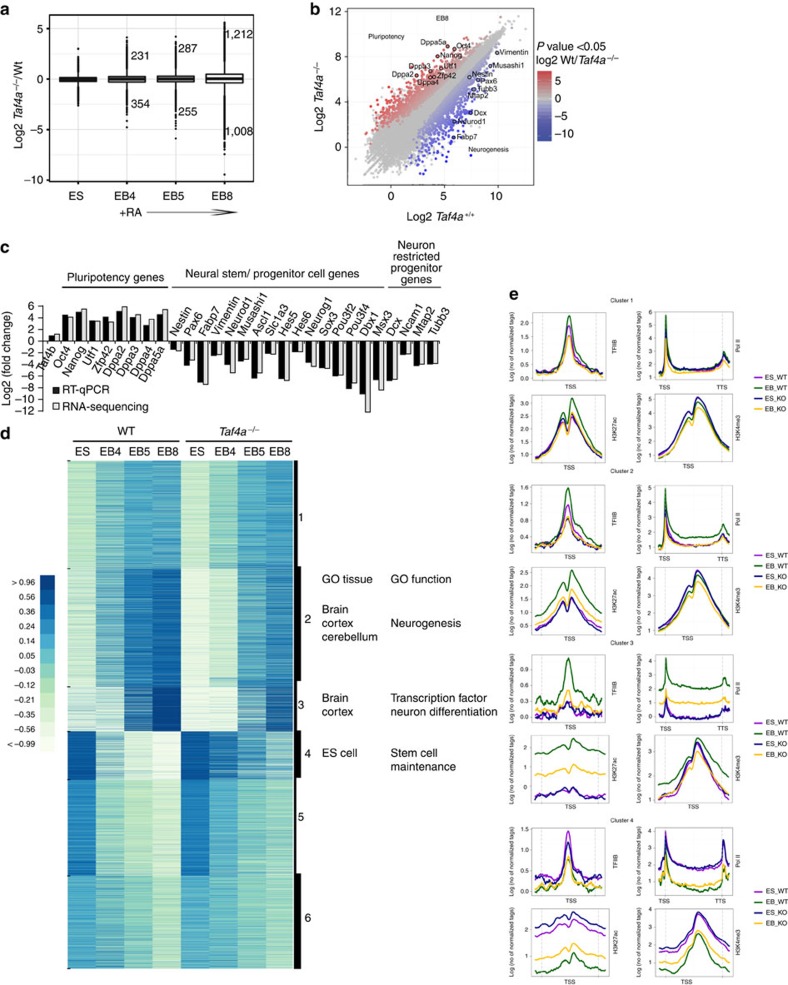
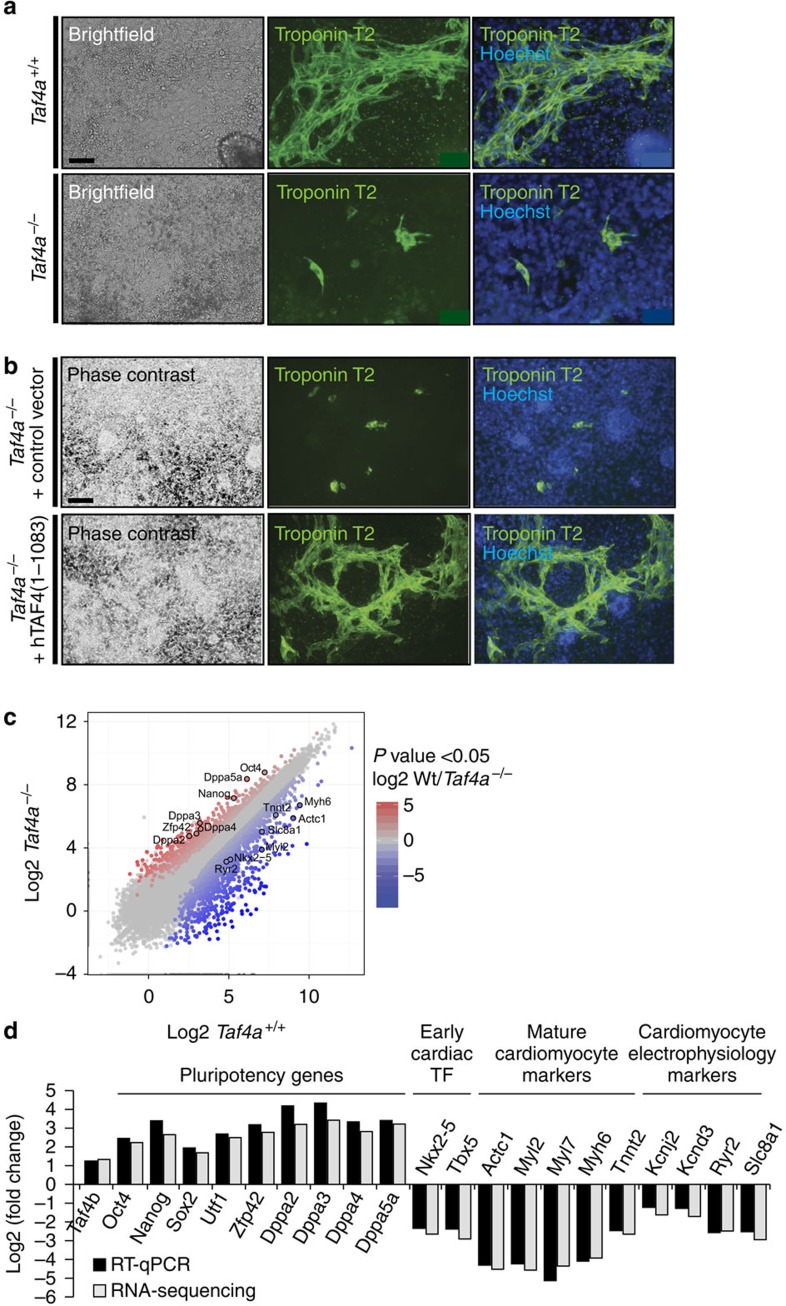
TAF4 (TATA-binding protein-associated factor 4) and its paralogue TAF4b are components of the TFIID core module. We inactivated the murine Taf4a gene to address Taf4 function during embryogenesis. Here we show that Taf4a(-/-) embryos survive until E9.5 where primary germ layers and many embryonic structures are identified showing Taf4 is dispensable for their specification. In contrast, Taf4 is required for correct patterning of the trunk and anterior structures, ventral morphogenesis and proper heart positioning. Overlapping expression of Taf4a and Taf4b during embryogenesis suggests their redundancy at early stages. In agreement with this, Taf4a(-/-) embryonic stem cells (ESCs) are viable and comprise Taf4b-containing TFIID. Nevertheless, Taf4a(-/-) ESCs do not complete differentiation into glutamatergic neurons and cardiomyocytes in vitro due to impaired preinitiation complex formation at the promoters of critical differentiation genes. We define an essential role of a core TFIID TAF in differentiation events during mammalian embryogenesis.
Figures
References
-
- Bieniossek C. et al. The architecture of human general transcription factor TFIID core complex. Nature 493, 699–702 (2013) . - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
