

Review
doi: 10.1038/ncb3330.
The ever-expanding role of HIF in tumour and stromal biology
Affiliations
- PMID: 27027486
- PMCID: PMC4898054
- DOI: 10.1038/ncb3330
Item in Clipboard
Review
The ever-expanding role of HIF in tumour and stromal biology
Nat Cell Biol.
2016 Apr.
Abstract
Low oxygen tension (hypoxia) is a hallmark of cancer that influences cancer cell function, but is also an important component of the tumour microenvironment as it alters the extracellular matrix, modulates the tumour immune response and increases angiogenesis. Here we discuss the regulation and role of hypoxia and its key transcriptional mediators, the hypoxia-inducible factor (HIF) family of transcription factors, in the tumour microenvironment and stromal compartments.
Figures
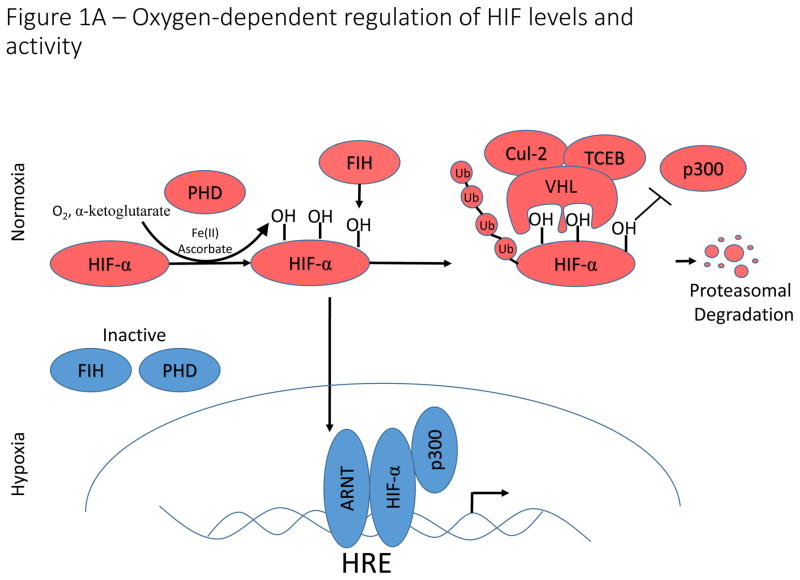
A)
Oxygen-dependent regulation of HIF-1α/HIF-2α. Under conditions in which sufficient oxygen is present (normoxia), prolyl hydroxylases (PHD) catalyze the hydroxylation of two key proline residues within the oxygen dependent degradation domains of HIF-1α and HIF-2α. The hydroxylation reaction catalyzed by PHDs utilizes molecular oxygen and α-ketoglutarate (which is converted to succinate in the reaction) as co-substrates and ferrous iron (Fe(II)) and ascorbate as cofactors. These hydroxylation events form a binding site for the E3 ubiquitin ligase, VHL, which catalyzes ubiquitination and subsequent proteasomal degradation. An additional asparagine residue in the C-terminal activation domain is hydroxylated by factor inhibiting HIF (FIH). Hydroxylation of this asparagine residue disrupts binding of p300 to HIF-α, thereby inhibiting HIF-transcriptional activation potential. Under hypoxic conditions, PHD and FIH activity is inhibited and unhydroxylated HIF-1α and HIF-2α translocate to the nucleus, form a complex with ARNT and p300, and activate transcription of HIF-target genes. B).
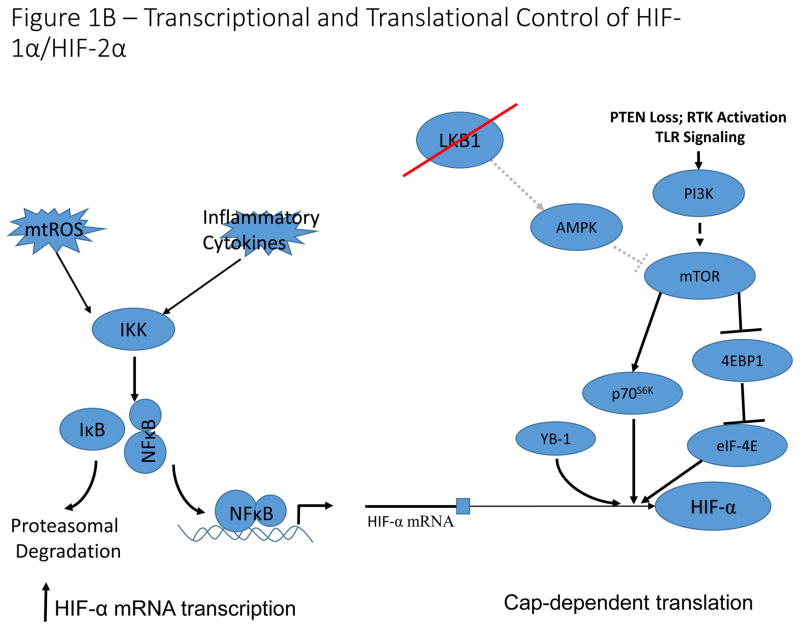
Transcriptional and translational control of HIF-1α/HIF-2α expression . Reactive oxygen species and pro-inflammatory conditions in the tumour microenvironment stimulate NF-κB-dependent transcriptional activation of HIF-1α, leading to increased expression under normoxic conditions. Activation of the mTOR signaling pathway by PI3K activation or LKB1 loss of function, results in increased cap-dependent translation of HIF-α mRNA. Additionally, the RNA/DNA binding protein, YB-1 is induced in multiple cancer types, and can bind to HIF-1α mRNA to stimulate cap-dependent translation. This leads to increased normoxic expression of HIF- α in cells with constitutively active PI3K signaling.
A)
Oxygen-dependent regulation of HIF-1α/HIF-2α. Under conditions in which sufficient oxygen is present (normoxia), prolyl hydroxylases (PHD) catalyze the hydroxylation of two key proline residues within the oxygen dependent degradation domains of HIF-1α and HIF-2α. The hydroxylation reaction catalyzed by PHDs utilizes molecular oxygen and α-ketoglutarate (which is converted to succinate in the reaction) as co-substrates and ferrous iron (Fe(II)) and ascorbate as cofactors. These hydroxylation events form a binding site for the E3 ubiquitin ligase, VHL, which catalyzes ubiquitination and subsequent proteasomal degradation. An additional asparagine residue in the C-terminal activation domain is hydroxylated by factor inhibiting HIF (FIH). Hydroxylation of this asparagine residue disrupts binding of p300 to HIF-α, thereby inhibiting HIF-transcriptional activation potential. Under hypoxic conditions, PHD and FIH activity is inhibited and unhydroxylated HIF-1α and HIF-2α translocate to the nucleus, form a complex with ARNT and p300, and activate transcription of HIF-target genes. B).
Transcriptional and translational control of HIF-1α/HIF-2α expression . Reactive oxygen species and pro-inflammatory conditions in the tumour microenvironment stimulate NF-κB-dependent transcriptional activation of HIF-1α, leading to increased expression under normoxic conditions. Activation of the mTOR signaling pathway by PI3K activation or LKB1 loss of function, results in increased cap-dependent translation of HIF-α mRNA. Additionally, the RNA/DNA binding protein, YB-1 is induced in multiple cancer types, and can bind to HIF-1α mRNA to stimulate cap-dependent translation. This leads to increased normoxic expression of HIF- α in cells with constitutively active PI3K signaling.
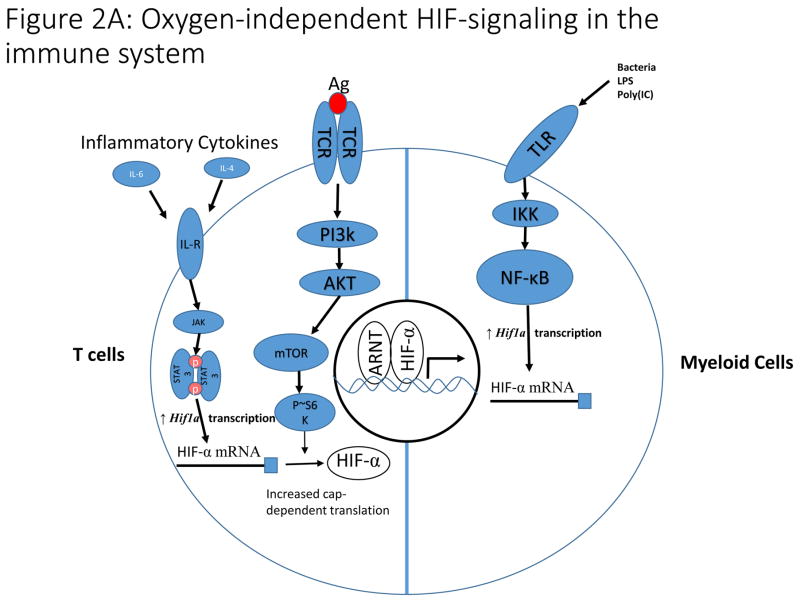
A). In T cells, antigen-dependent activation of the T cell Receptor (TCR) signals through the PI3K pathway to stimulate increased translation of HIF-α mRNA. Additionally, inflammatory cytokines signal through the JAK/STAT3 pathway to increase transcription of HIF-α mRNA. These pathways are essential for induction of HIF-signaling in hypoxia and during T cell activation. In myeloid cells, toll-like receptor (TLR) signaling increases NF-κB-dependent transcription of HIF-α mRNA in response to a variety of stimuli including bacterial infection leading to increased HIF-signaling. B). The hydroxylation reaction catalyzed by PHDs utilizes α-ketoglutarate as a co-substrate, producing succinate in the process. Loss of function mutations to succinate dehydrogenase (SDH) and fumarate hydratase (FH) result in accumulation of succinate, which inhibits PHD activity and stabilizes HIF-α in normoxia. The enzymes labelled in red (SDH, FH, and IDH) are frequently mutated in a variety of malignancies resulting in inhibition of PHD activity and stabilization of HIF-α subunits. Increased concentrations of lactate and pyruvate have also been demonstrated to promote stabilization of HIF-α, but the mechanism by which this stabilization occurs remains unclear.
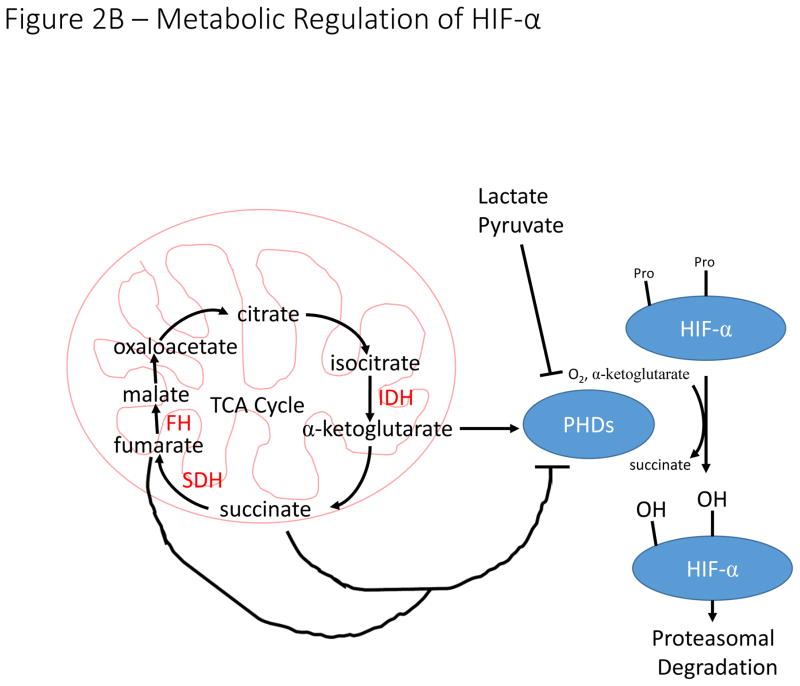
A). In T cells, antigen-dependent activation of the T cell Receptor (TCR) signals through the PI3K pathway to stimulate increased translation of HIF-α mRNA. Additionally, inflammatory cytokines signal through the JAK/STAT3 pathway to increase transcription of HIF-α mRNA. These pathways are essential for induction of HIF-signaling in hypoxia and during T cell activation. In myeloid cells, toll-like receptor (TLR) signaling increases NF-κB-dependent transcription of HIF-α mRNA in response to a variety of stimuli including bacterial infection leading to increased HIF-signaling. B). The hydroxylation reaction catalyzed by PHDs utilizes α-ketoglutarate as a co-substrate, producing succinate in the process. Loss of function mutations to succinate dehydrogenase (SDH) and fumarate hydratase (FH) result in accumulation of succinate, which inhibits PHD activity and stabilizes HIF-α in normoxia. The enzymes labelled in red (SDH, FH, and IDH) are frequently mutated in a variety of malignancies resulting in inhibition of PHD activity and stabilization of HIF-α subunits. Increased concentrations of lactate and pyruvate have also been demonstrated to promote stabilization of HIF-α, but the mechanism by which this stabilization occurs remains unclear.
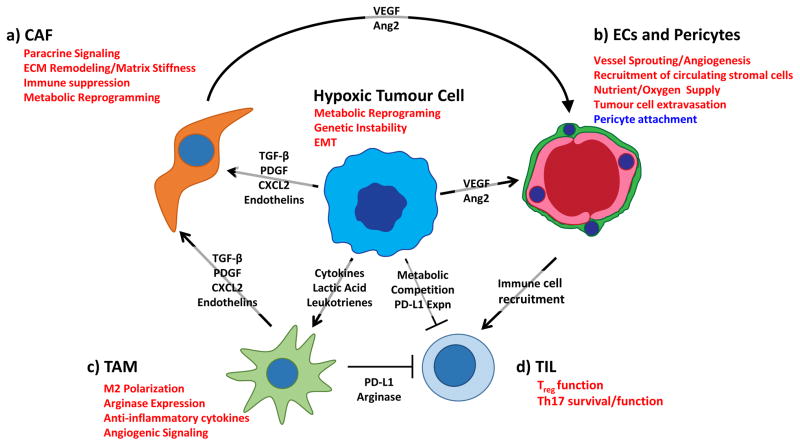
Hypoxic tumour and stromal cells initiate paracrine signalling to stimulate angiogenesis and recruit stromal cells from the circulation, creating an immunosuppressive microenvironment. A). Hypoxia stimulates tumour cells and TAMs to secrete paracrine factors (TGF-β, PDGF, CXCL2, Endothelin) that promote activation of cancer-associated fibroblasts. In CAFs, hypoxia stimulates extracellular matrix (ECM) remodelling, which promotes increased tumour aggressiveness through stiffening of the ECM. Hypoxic CAFs also synthesize and release factors that drive angiogenesis and immune cell recruitment to the tumour site. B). Hypoxia drives tumour and stromal secretion of vascular endothelial growth factor (VEGF) and other pro-angiogenic factors that recruit endothelial cells and pericytes from the surrounding vasculature. Hypoxia directly effects vascular barrier function by decreasing the association between pericytes and endothelial cells, thereby facilitating tumour cell extravasation and recruitment of stromal cells from the circulation. C). Hypoxia stimulates recruitment of circulating macrophages and promotes alternative (M2) activation by increasing expression of macrophage chemotractants and lactate levels. Hypoxic M2 macrophages create a functionally immunosuppressive microenvironment by increasing expression of arginase and immune checkpoint ligands. D). Hypoxia promotes a functionally immunosuppressive microenvironment by stimulating Treg cell function and increasing expression of immune checkpoint molecules such as PD-L1 and CTLA4 on tumour cells. Hypoxia also drives metabolic reprogramming in tumour cells, allowing them to out-compete T cells for key metabolites critical for T cell function. Hypoxia prolongs the survival of TH17 cells, though the role of this helper T cell subtype in tumourigenesis remains controversial. Pathways in red are induced in hypoxia, while pathways in blue are suppressed under hypoxic conditions. Ang2, angiopoietin 2; CAF, cancer associated fibroblast; CTL, cytotoxic T cell; EC, endothelial cell; EMT, epithelial to mesenchymal transition; TAM, tumour-associated macrophage.
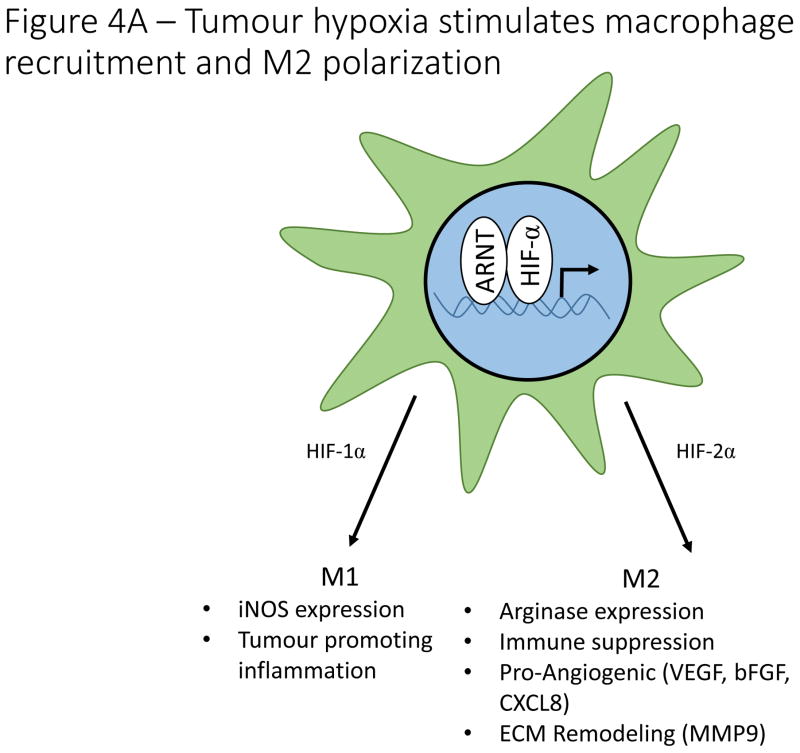
A). Hypoxia regulates macrophage polarization by controlling expression of genes involved in the function of M1 and M2 macrophages. HIF-1α promotes expression of the M1 gene, iNOS, while HIF-2α promotes expression of the M2 gene, Arg1. Hypoxia induces macrophage production of genes involved in angiogenesis (VEGF, bFGF, CXCL12), ECM remodeling (MMP9), and immune suppression (Arg1). B). HIF signaling plays a controversial role in determining the differentiation of naïve CD4+ T cells into either Th17 pro-inflammatory T cells or FoxP3+ T reg cells with reports suggesting that HIF either induces or inhibits the formation of both cell types. In contrast, overwhelming evidence indicates that hypoxia stimulates the secretion of a number of cytokines and chemoattractants from cancer cells and tumour associated macrophages that recruit T regs from the circulation. On a cell intrinsic basis, hypoxia stimulates T reg production of CD73, thereby increasing adenosine levels in the tumour microenvironment, resulting in inhibition of effector T cells. Hypoxia also stimulates T reg production of other immune suppressive molecules, including Galectin-1, TGF-β and IL-10.
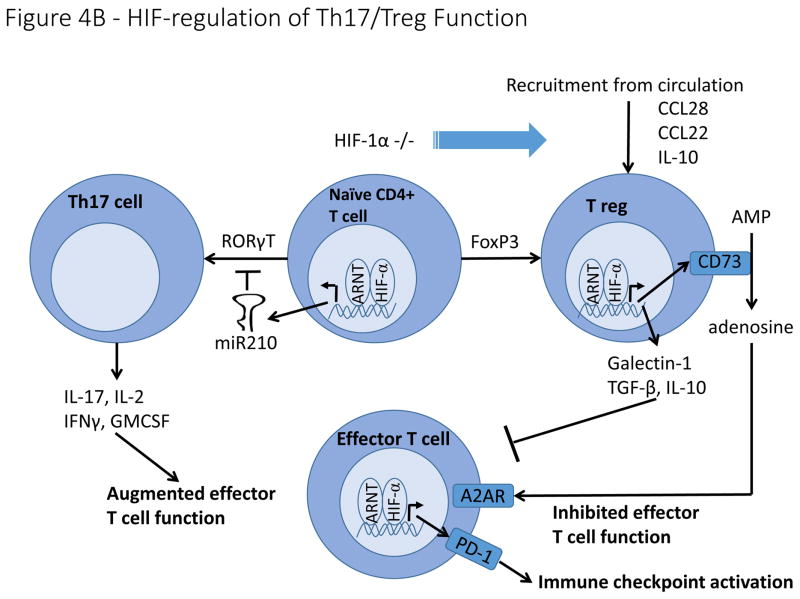
A). Hypoxia regulates macrophage polarization by controlling expression of genes involved in the function of M1 and M2 macrophages. HIF-1α promotes expression of the M1 gene, iNOS, while HIF-2α promotes expression of the M2 gene, Arg1. Hypoxia induces macrophage production of genes involved in angiogenesis (VEGF, bFGF, CXCL12), ECM remodeling (MMP9), and immune suppression (Arg1). B). HIF signaling plays a controversial role in determining the differentiation of naïve CD4+ T cells into either Th17 pro-inflammatory T cells or FoxP3+ T reg cells with reports suggesting that HIF either induces or inhibits the formation of both cell types. In contrast, overwhelming evidence indicates that hypoxia stimulates the secretion of a number of cytokines and chemoattractants from cancer cells and tumour associated macrophages that recruit T regs from the circulation. On a cell intrinsic basis, hypoxia stimulates T reg production of CD73, thereby increasing adenosine levels in the tumour microenvironment, resulting in inhibition of effector T cells. Hypoxia also stimulates T reg production of other immune suppressive molecules, including Galectin-1, TGF-β and IL-10.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
