

Spindle Bursts in Neonatal Rat Cerebral Cortex
- PMID: 27034844
- PMCID: PMC4806652
- DOI: 10.1155/2016/3467832
Spindle Bursts in Neonatal Rat Cerebral Cortex
Abstract
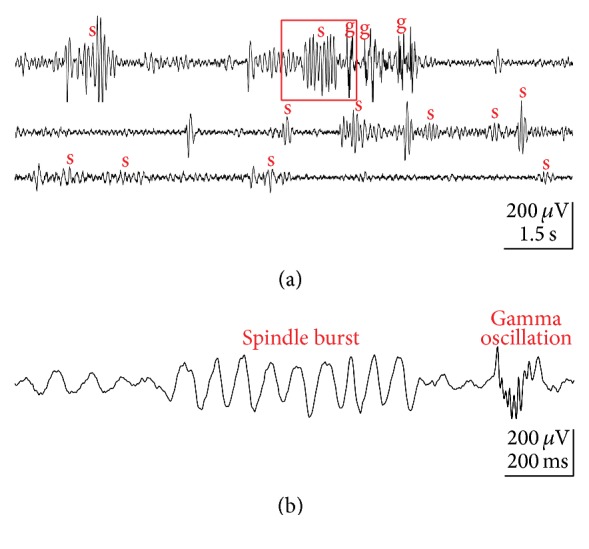
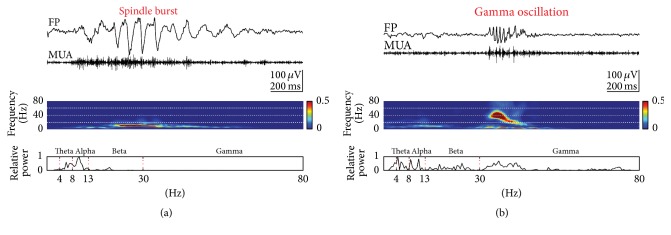
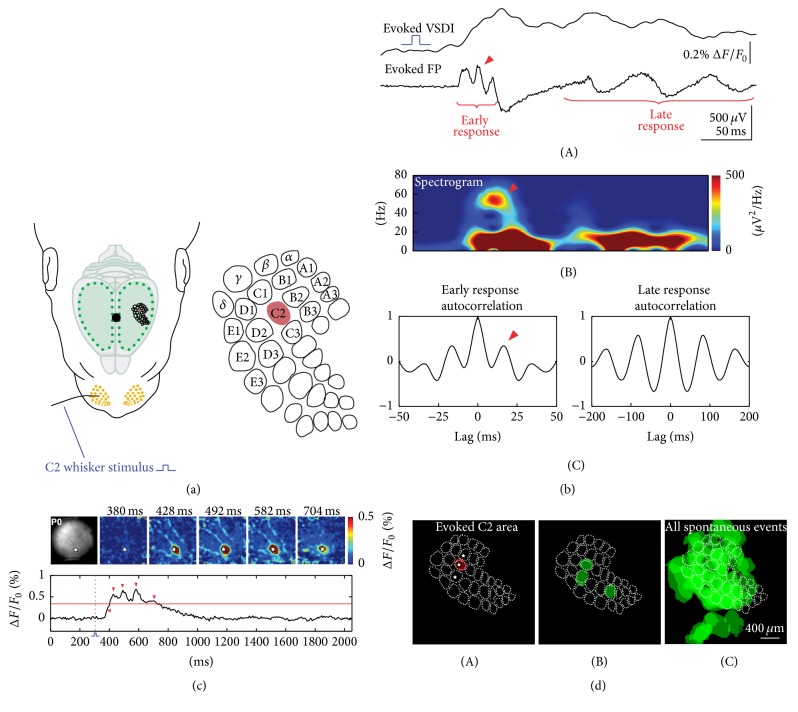
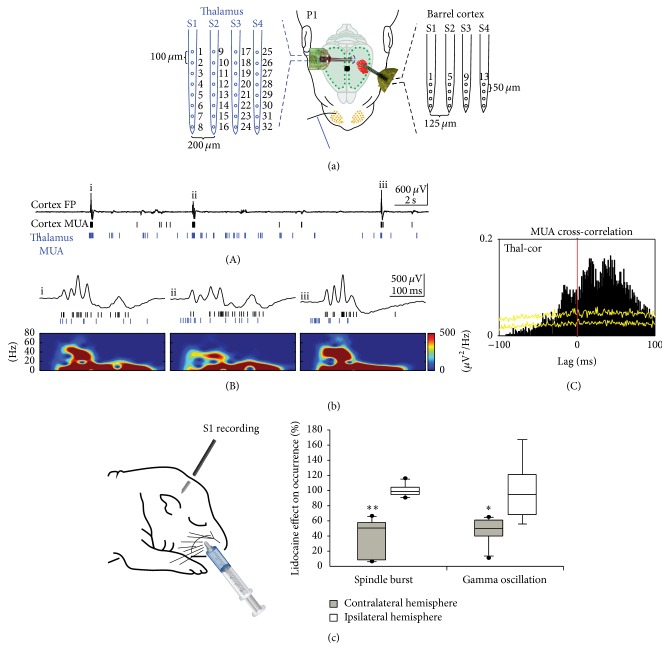
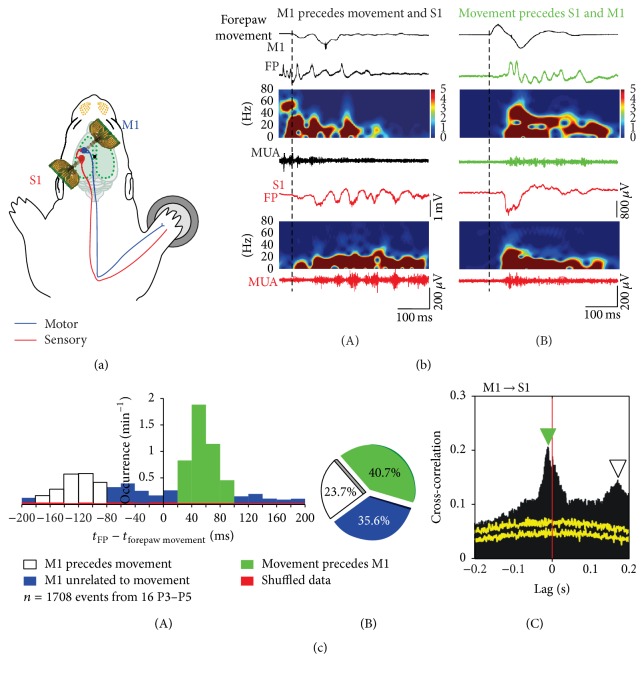
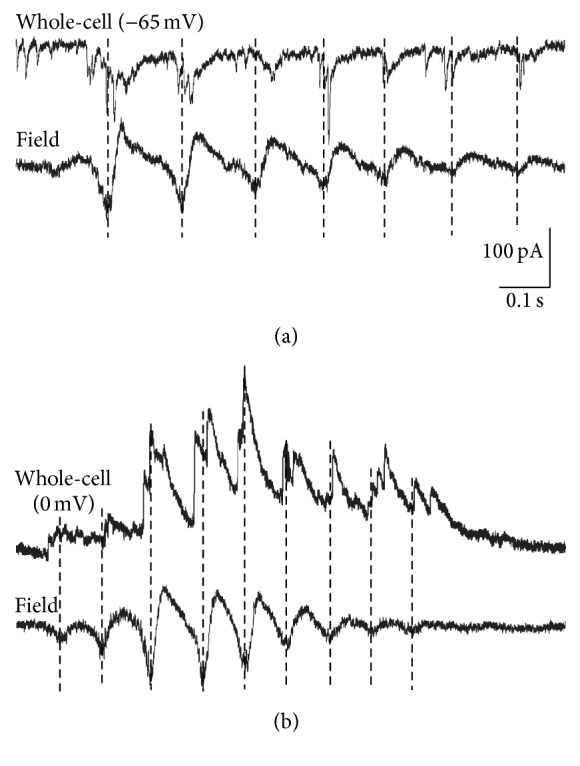
Spontaneous and sensory evoked spindle bursts represent a functional hallmark of the developing cerebral cortex in vitro and in vivo. They have been observed in various neocortical areas of numerous species, including newborn rodents and preterm human infants. Spindle bursts are generated in complex neocortical-subcortical circuits involving in many cases the participation of motor brain regions. Together with early gamma oscillations, spindle bursts synchronize the activity of a local neuronal network organized in a cortical column. Disturbances in spindle burst activity during corticogenesis may contribute to disorders in cortical architecture and in the activity-dependent control of programmed cell death. In this review we discuss (i) the functional properties of spindle bursts, (ii) the mechanisms underlying their generation, (iii) the synchronous patterns and cortical networks associated with spindle bursts, and (iv) the physiological and pathophysiological role of spindle bursts during early cortical development.
Figures
Similar articles
-
Neuronal activity patterns in the developing barrel cortex.Neuroscience. 2018 Jan 1;368:256-267. doi: 10.1016/j.neuroscience.2017.05.025. Epub 2017 May 19. Neuroscience. 2018. PMID: 28528963 Review.
-
Sensory-evoked and spontaneous gamma and spindle bursts in neonatal rat motor cortex.J Neurosci. 2014 Aug 13;34(33):10870-83. doi: 10.1523/JNEUROSCI.4539-13.2014. J Neurosci. 2014. PMID: 25122889 Free PMC article.
-
Voltage-sensitive dye imaging reveals dynamic spatiotemporal properties of cortical activity after spontaneous muscle twitches in the newborn rat.J Neurosci. 2012 Aug 8;32(32):10982-94. doi: 10.1523/JNEUROSCI.1322-12.2012. J Neurosci. 2012. PMID: 22875932 Free PMC article.
-
Three patterns of oscillatory activity differentially synchronize developing neocortical networks in vivo.J Neurosci. 2009 Jul 15;29(28):9011-25. doi: 10.1523/JNEUROSCI.5646-08.2009. J Neurosci. 2009. PMID: 19605639 Free PMC article.
-
Early patterns of electrical activity in the developing cerebral cortex of humans and rodents.Trends Neurosci. 2006 Jul;29(7):414-418. doi: 10.1016/j.tins.2006.05.007. Epub 2006 May 19. Trends Neurosci. 2006. PMID: 16713634 Review.
Cited by
-
Electrical activity controls area-specific expression of neuronal apoptosis in the mouse developing cerebral cortex.Elife. 2017 Aug 21;6:e27696. doi: 10.7554/eLife.27696. Elife. 2017. PMID: 28826501 Free PMC article.
-
Micro Three-Dimensional Neuronal Cultures Generate Developing Cortex-Like Activity Patterns.Front Neurosci. 2020 Oct 2;14:563905. doi: 10.3389/fnins.2020.563905. eCollection 2020. Front Neurosci. 2020. PMID: 33122989 Free PMC article.
-
Reelin Regulates Developmental Desynchronization Transition of Neocortical Network Activity.Biomolecules. 2024 May 17;14(5):593. doi: 10.3390/biom14050593. Biomolecules. 2024. PMID: 38786001 Free PMC article.
-
Spontaneous Neuronal Activity in Developing Neocortical Networks: From Single Cells to Large-Scale Interactions.Front Neural Circuits. 2016 May 24;10:40. doi: 10.3389/fncir.2016.00040. eCollection 2016. Front Neural Circuits. 2016. PMID: 27252626 Free PMC article. Review.
-
Sleep as a window on the sensorimotor foundations of the developing hippocampus.Hippocampus. 2022 Feb;32(2):89-97. doi: 10.1002/hipo.23334. Epub 2021 May 4. Hippocampus. 2022. PMID: 33945190 Free PMC article. Review.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
