

The Genetic Cost of Neanderthal Introgression
- PMID: 27038113
- PMCID: PMC4896200
- DOI: 10.1534/genetics.116.186890
The Genetic Cost of Neanderthal Introgression
Abstract
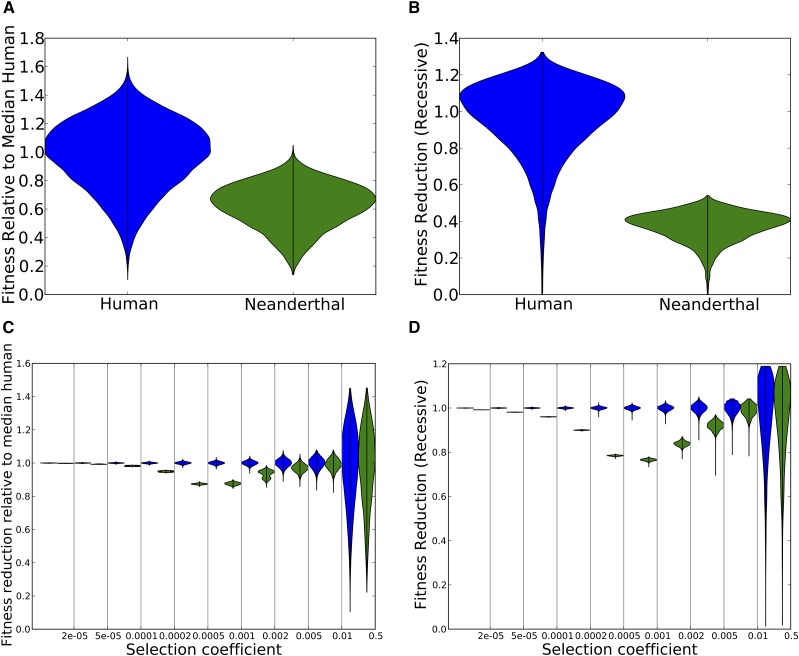
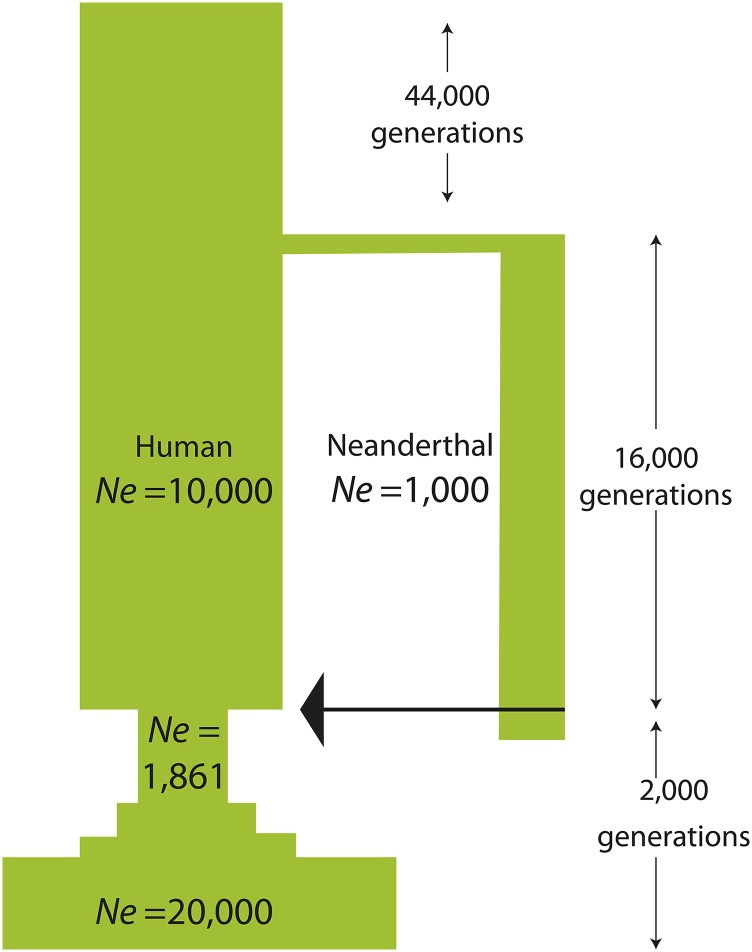
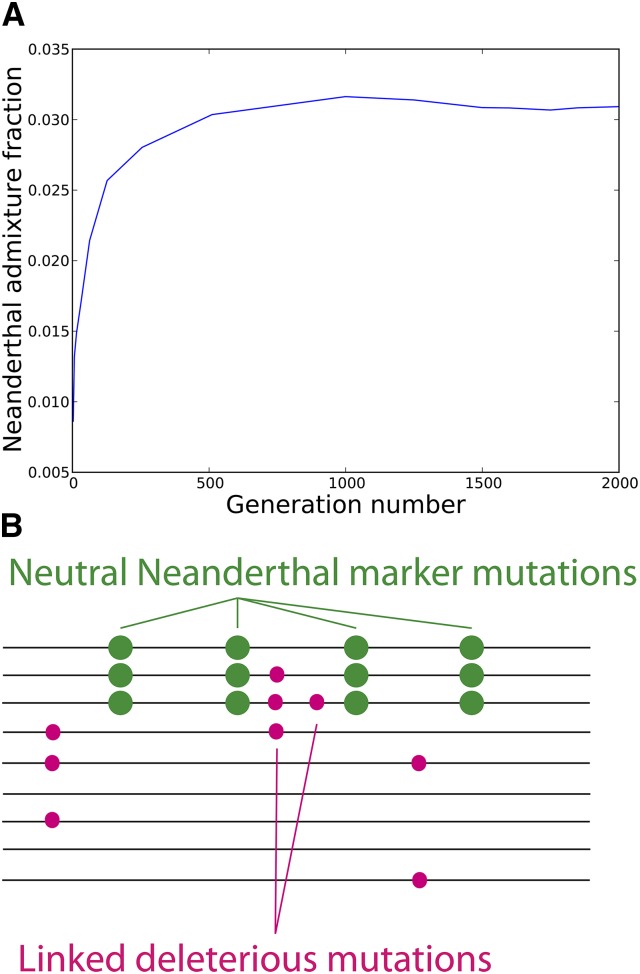
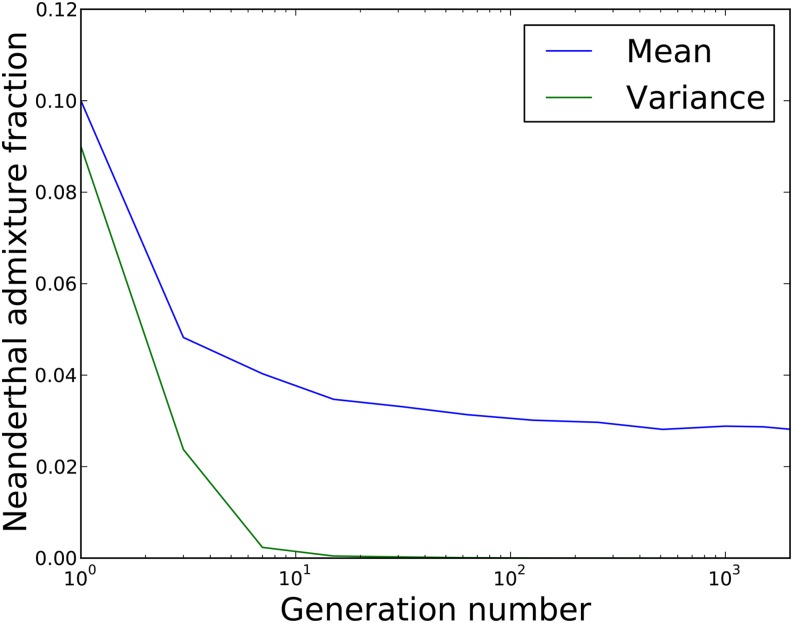
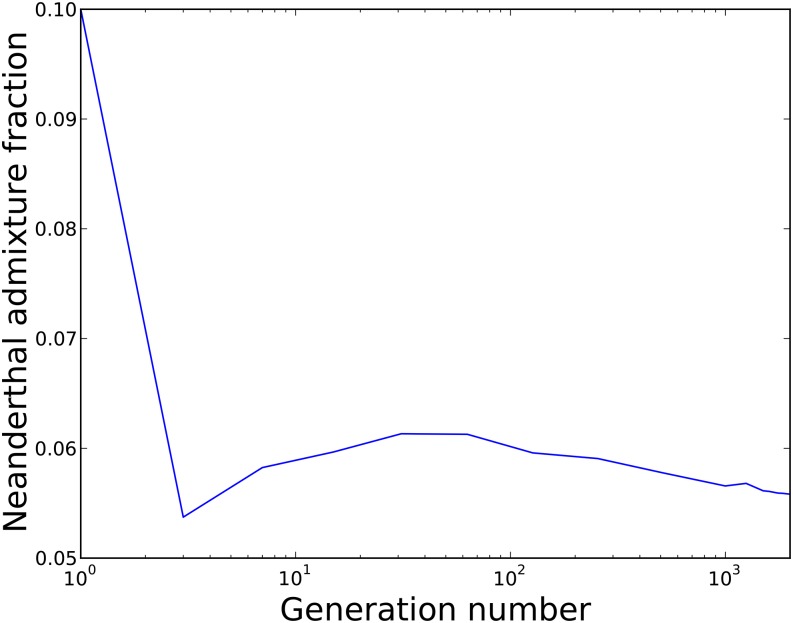
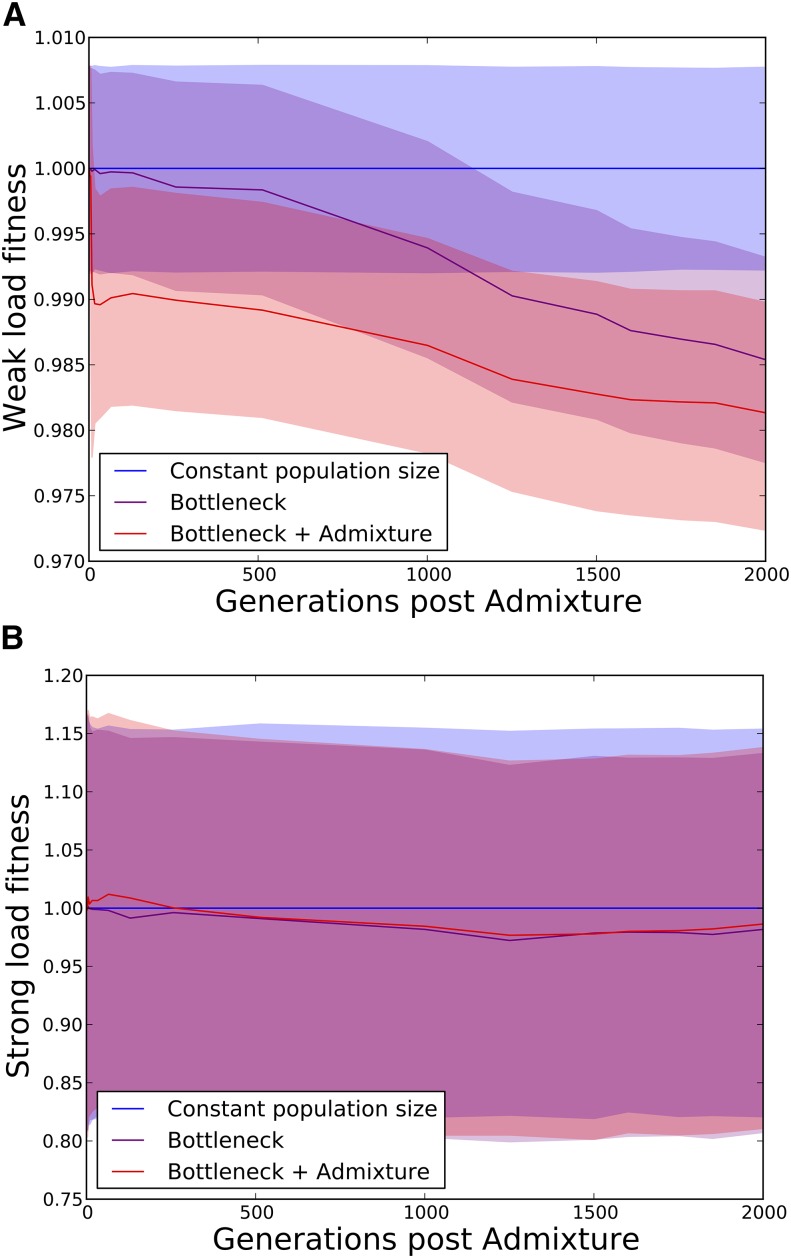
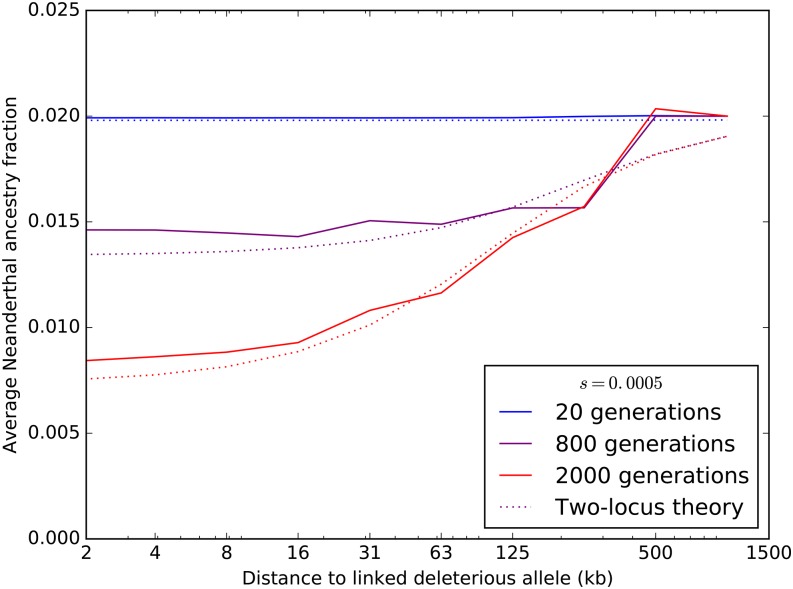
Approximately 2-4% of genetic material in human populations outside Africa is derived from Neanderthals who interbred with anatomically modern humans. Recent studies have shown that this Neanderthal DNA is depleted around functional genomic regions; this has been suggested to be a consequence of harmful epistatic interactions between human and Neanderthal alleles. However, using published estimates of Neanderthal inbreeding and the distribution of mutational fitness effects, we infer that Neanderthals had at least 40% lower fitness than humans on average; this increased load predicts the reduction in Neanderthal introgression around genes without the need to invoke epistasis. We also predict a residual Neanderthal mutational load in non-Africans, leading to a fitness reduction of at least 0.5%. This effect of Neanderthal admixture has been left out of previous debate on mutation load differences between Africans and non-Africans. We also show that if many deleterious mutations are recessive, the Neanderthal admixture fraction could increase over time due to the protective effect of Neanderthal haplotypes against deleterious alleles that arose recently in the human population. This might partially explain why so many organisms retain gene flow from other species and appear to derive adaptive benefits from introgression.
Keywords: archaic hominins; deleterious mutation load; gene flow; heterosis; nearly neutral theory.
Copyright © 2016 by the Genetics Society of America.
Figures
References
-
- Allendorf F., Hohenlohe P., Luikart G., 2010. Genomics and the future of conservation genetics. Nat. Rev. Genet. 11: 697–709. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
