

The Functional Role of eL19 and eB12 Intersubunit Bridge in the Eukaryotic Ribosome
- PMID: 27038511
- PMCID: PMC4884501
- DOI: 10.1016/j.jmb.2016.03.023
The Functional Role of eL19 and eB12 Intersubunit Bridge in the Eukaryotic Ribosome
Abstract
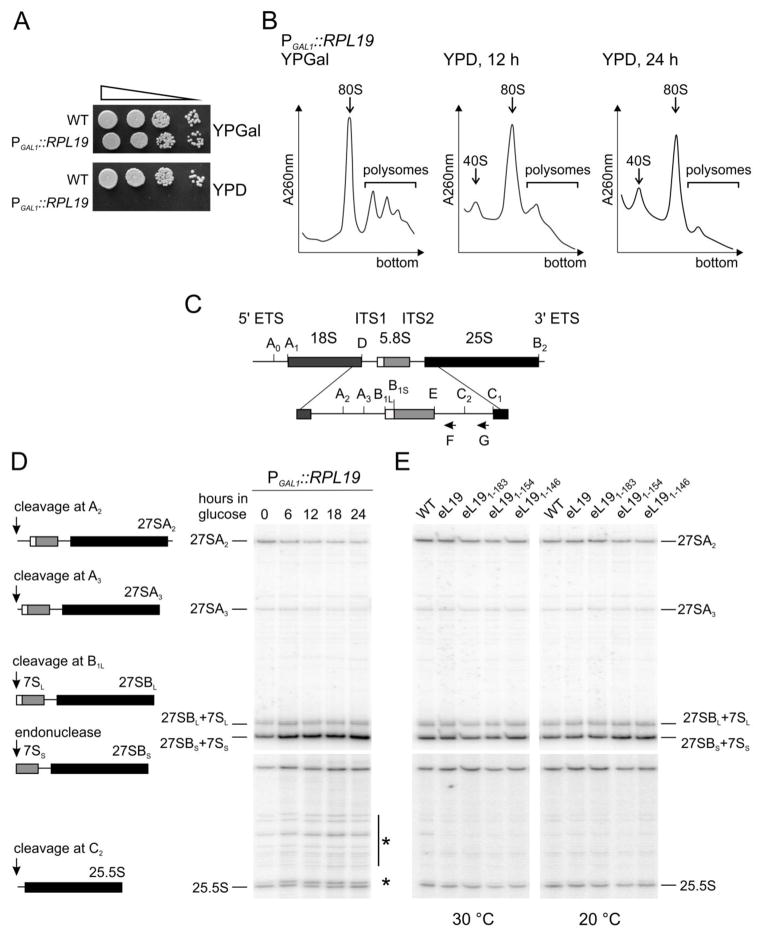
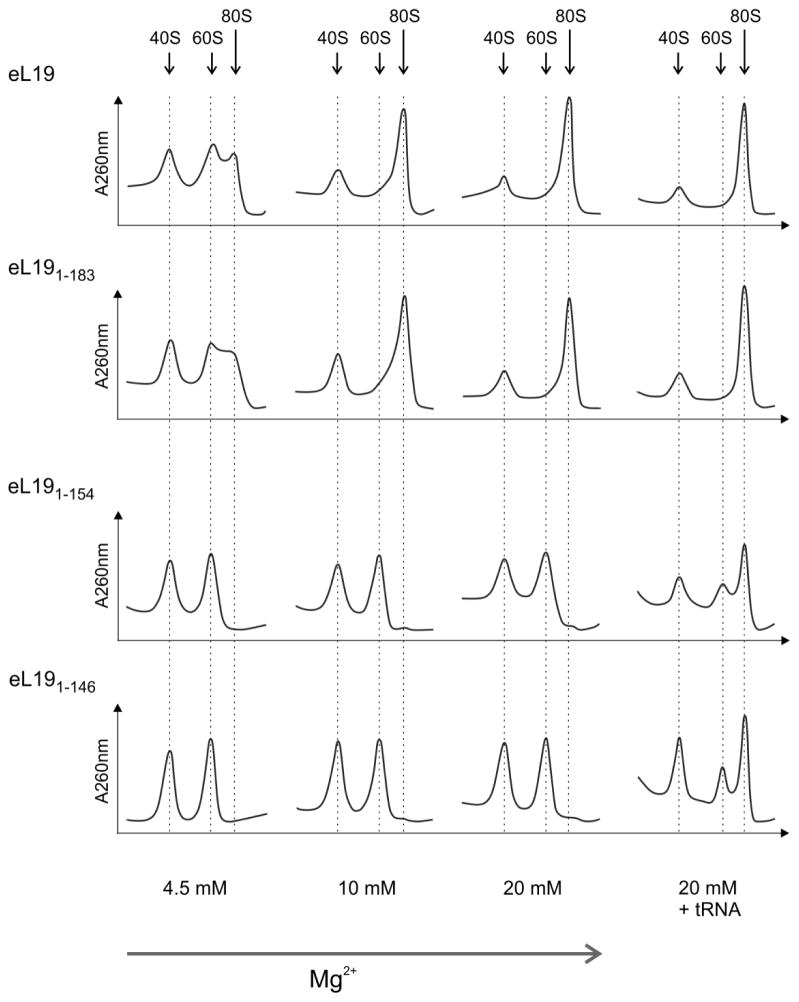
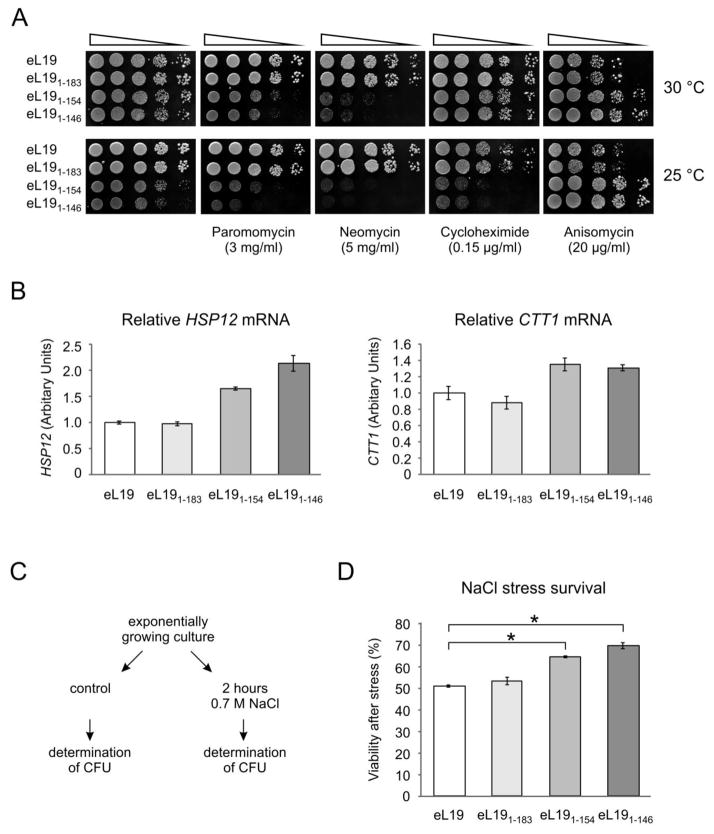
During translation, the two eukaryotic ribosomal subunits remain associated through 17 intersubunit bridges, five of which are eukaryote specific. These are mainly localized to the peripheral regions and are believed to stabilize the structure of the ribosome. The functional importance of these bridges remains largely unknown. Here, the essentiality of the eukaryote-specific bridge eB12 has been investigated. The main component of this bridge is ribosomal protein eL19 that is composed of an N-terminal globular domain, a middle region, and a long C-terminal α-helix. The analysis of deletion mutants demonstrated that the globular domain and middle region of eL19 are essential for cell viability, most likely functioning in ribosome assembly. The eB12 bridge, formed by contacts between the C-terminal α-helix of eL19 and 18S rRNA in concert with additional stabilizing interactions involving either eS7 or uS17, is dispensable for viability. Nevertheless, eL19 mutants impaired in eB12 bridge formation displayed slow growth phenotypes, altered sensitivity/resistance to translational inhibitors, and enhanced hyperosmotic stress tolerance. Biochemical analyses determined that the eB12 bridge contributes to the stability of ribosome subunit interactions in vitro. 60S subunits containing eL19 variants defective in eB12 bridge formation failed to form 80S ribosomes regardless of Mg(2+) concentration. The reassociation of 40S and mutant 60S subunits was markedly improved in the presence of deacetylated tRNA, emphasizing the importance of tRNAs during the subunit association. We propose that the eB12 bridge plays an important role in subunit joining and in optimizing ribosome functionality.
Keywords: Saccharomyces cerevisiae; eukaryote-specific bridges; ribosome assembly; ribosome subunit association; stress tolerance.
Copyright © 2016 Elsevier Ltd. All rights reserved.
Figures


Similar articles
-
Ribosomal protein eL24, involved in two intersubunit bridges, stimulates translation initiation and elongation.Nucleic Acids Res. 2019 Jan 10;47(1):406-420. doi: 10.1093/nar/gky1083. Nucleic Acids Res. 2019. PMID: 30407570 Free PMC article.
-
Functional Interactions of Ribosomal Intersubunit Bridges in Saccharomyces cerevisiae.Genetics. 2019 Dec;213(4):1329-1339. doi: 10.1534/genetics.119.302777. Epub 2019 Oct 24. Genetics. 2019. PMID: 31649153 Free PMC article.
-
The Intersubunit Bridge B1b of the Bacterial Ribosome Facilitates Initiation of Protein Synthesis and Maintenance of Translational Fidelity.J Mol Biol. 2017 Apr 7;429(7):1067-1080. doi: 10.1016/j.jmb.2017.02.015. Epub 2017 Feb 24. J Mol Biol. 2017. PMID: 28238762
-
The structure and function of the eukaryotic ribosome.Cold Spring Harb Perspect Biol. 2012 May 1;4(5):a011536. doi: 10.1101/cshperspect.a011536. Cold Spring Harb Perspect Biol. 2012. PMID: 22550233 Free PMC article. Review.
-
'View From A Bridge': A New Perspective on Eukaryotic rRNA Base Modification.Trends Biochem Sci. 2015 Oct;40(10):560-575. doi: 10.1016/j.tibs.2015.07.008. Trends Biochem Sci. 2015. PMID: 26410597 Review.
Cited by
-
Putting It All Together: The Roles of Ribosomal Proteins in Nucleolar Stages of 60S Ribosomal Assembly in the Yeast Saccharomyces cerevisiae.Biomolecules. 2024 Aug 9;14(8):975. doi: 10.3390/biom14080975. Biomolecules. 2024. PMID: 39199362 Free PMC article. Review.
-
Loss of Conserved rRNA Modifications in the Peptidyl Transferase Center Leads to Diminished Protein Synthesis and Cell Growth in Budding Yeast.Int J Mol Sci. 2024 May 10;25(10):5194. doi: 10.3390/ijms25105194. Int J Mol Sci. 2024. PMID: 38791231 Free PMC article.
-
A widely applicable and cost-effective method for specific RNA-protein complex isolation.Sci Rep. 2023 Apr 27;13(1):6898. doi: 10.1038/s41598-023-34157-0. Sci Rep. 2023. PMID: 37106019 Free PMC article.
-
Archaea/eukaryote-specific ribosomal proteins - guardians of a complex structure.Comput Struct Biotechnol J. 2023 Jan 27;21:1249-1261. doi: 10.1016/j.csbj.2023.01.037. eCollection 2023. Comput Struct Biotechnol J. 2023. PMID: 36817958 Free PMC article. Review.
-
Ribosomal protein eL24, involved in two intersubunit bridges, stimulates translation initiation and elongation.Nucleic Acids Res. 2019 Jan 10;47(1):406-420. doi: 10.1093/nar/gky1083. Nucleic Acids Res. 2019. PMID: 30407570 Free PMC article.
References
-
- Ben-Shem A, Garreau de Loubresse N, Melnikov S, Jenner L, Yusupova G, Yusupov M. The structure of the eukaryotic ribosome at 3.0 A resolution. Science. 2011;334:1524–9. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
