

Review
doi: 10.1038/nrmicro.2016.26.
Epub 2016 Apr 4.
Splitsville: structural and functional insights into the dynamic bacterial Z ring
Affiliations
- PMID: 27040757
- PMCID: PMC5290750
- DOI: 10.1038/nrmicro.2016.26
Item in Clipboard
Review
Splitsville: structural and functional insights into the dynamic bacterial Z ring
Nat Rev Microbiol.
2016 Apr.
Abstract
Bacteria must divide to increase in number and colonize their niche. Binary fission is the most widespread means of bacterial cell division, but even this relatively simple mechanism has many variations on a theme. In most bacteria, the tubulin homologue FtsZ assembles into a ring structure, termed the Z ring, at the site of cytokinesis and recruits additional proteins to form a large protein machine - the divisome - that spans the membrane. In this Review, we discuss current insights into the regulation of the assembly of the Z ring and how the divisome drives membrane invagination and septal cell wall growth while flexibly responding to various cellular inputs.
Figures
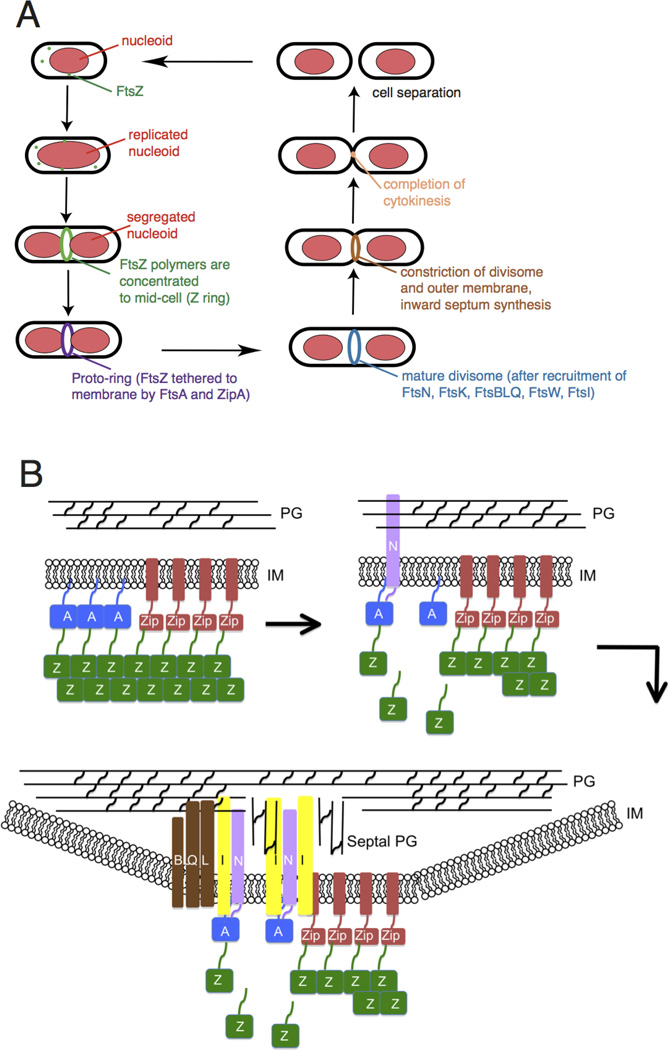
Overview of bacterial cytokinesis. (A) After E. coli cells replicate and segregate their chromosomes, organized as nucleoids, FtsZ and its membrane tethers are concentrated at midcell and organize into a Z ring or proto-ring. After recruitment of additional proteins, the proto-ring progresses into a mature divisome, which coordinates constriction of the inner and outer membranes with targeted cell wall hydrolases and ingrowing septal peptidoglycan. The ultimate result is separation into two equal-sized daughter cells. (B) ZipA and FtsA tether FtsZ protofilaments and polymer bundles to the membrane using flexible linkers and either a transmembrane segment (ZipA, red rectangle) or an amphipathic helix (FtsA). ZipA and FtsA are depicted as clustered, but they may be interspersed instead. In a later stage, ZipA bundling of FtsZ protofilaments is offset by FtsA-mediated disassembly. In addition, FtsA recruits FtsN via its cytoplasmic domain, which in turn may disrupt FtsA oligomers. FtsN’s periplasmic domain is targeted to peptidoglycan, reinforcing its localization and potentially stimulating FtsI and septal peptidoglycan synthesis. After septal peptidoglycan synthesis is underway, the other divisome proteins including the FtsBLQ complex transduce signals from the periplasm to the Z ring via FtsA, coordinating Z ring constriction (and FtsZ turnover) with septal peptidoglycan synthesis and targeted cell wall hydrolysis. For simplicity, many of the divisome proteins are not shown here, but they are shown in Fig. 4A.
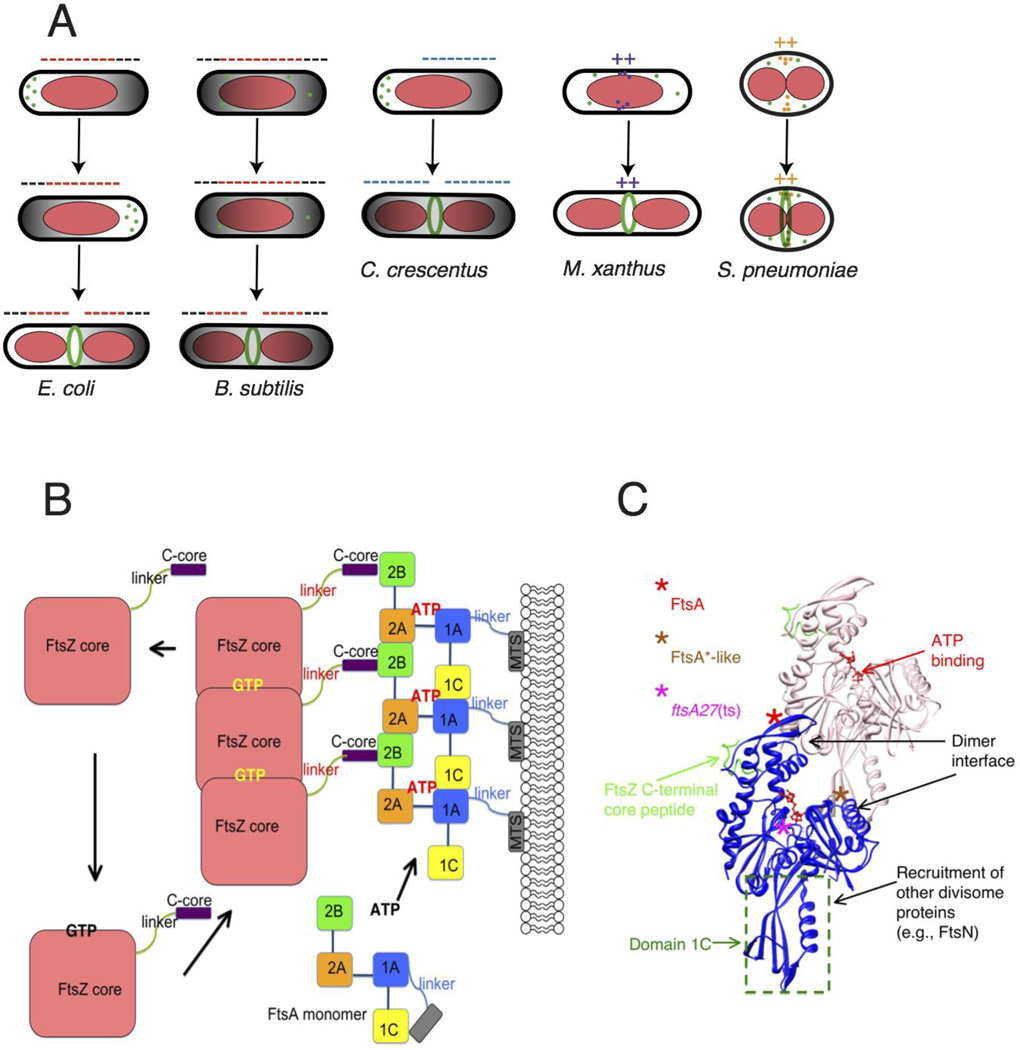
Placement and tethering of the Z ring. (A) The Z ring is targeted to the division site by related but distinct mechanisms. In E. coli and B. subtilis, a bipolar gradient formed by the Min system (black; oscillating in E. coli and fixed in B. subtilis) and nucleoid occlusion (red) inhibit assembly of FtsZ (minus signs) so that the Z ring is restricted to the lowest region of negative regulation at midcell. In C. crescentus, MipZ forms a bipolar gradient in response to cell cycle cues and negatively regulates Z ring positioning. In contrast, FtsZ assembly stimulators in M. xanthus and S. pneumoniae localize at midcell via unknown mechanisms, potentiating assembly of the Z ring there (plus signs). (B) GTP promotes assembly of FtsZ into protofilaments, which attach to subdomain 2B of FtsA via FtsZ’s C-terminal core at the terminus of a flexible linker. GTP hydrolysis is coupled to disassembly of FtsZ, as the cycle repeats. FtsA’s four subdomains are schematized, along with the amphipathic C-terminal membrane targeting sequence (MTS) at the end of a short linker. ATP binding promotes oligomerization of S. pneumoniae FtsA, but is not yet confirmed for E. coli FtsA. Reprinted with permission from Ref. . (C) Structure of an FtsA dimer and its bound C-terminal core peptide of FtsZ based on the atomic structure from Thermotoga maritima
. The flexible linker and amphipathic helix are not shown. Loss of function lesions such as the thermosensitive FtsA12 allele map near the ATP binding site, whereas gain of function (FtsA*, FtsA*-like) generally map to the dimer interface. FtsA subdomains have distinct activities, which are outlined.
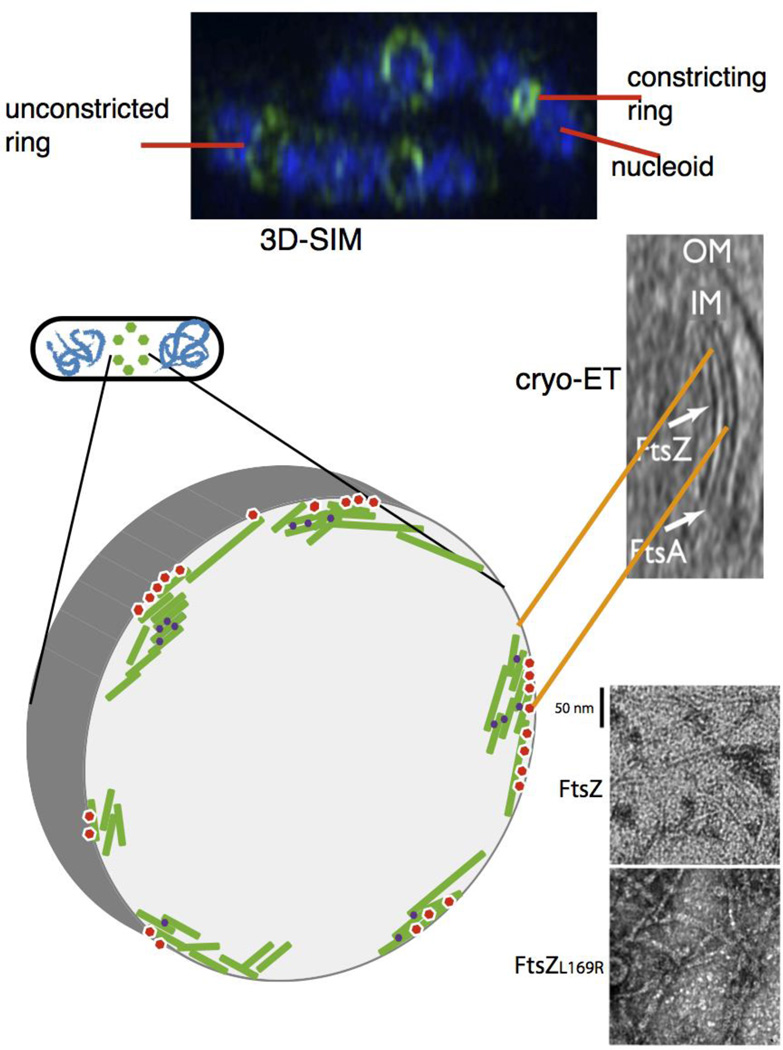
Proto-ring ultrastructure in E. coli. (A) 3D-SIM of several species indicates a patchy distribution of proto-ring proteins; shown is a 3D-SIM image of E. coli cells fixed and stained with DAPI (blue) for nucleoids and anti-FtsZ (green). Image courtesy of V. Rowlett. (B) the modeled structure based on this patchy pattern is depicted, with FtsZ protofilaments (green) attached to membrane tethers (red) and bundled by crosslinking Zap proteins (purple). (C) a cryo-ET image of E. coli producing FtsA and a mutant FtsZ that increases its stability, showing concentric filaments of both against the inner membrane (reproduced with permission from ref. ). This is also modeled in the schematic in (B). (D) transmission EM images of purified FtsZ or FtsZL169R mutant in the presence of GTP, showing the double protofilaments for FtsZL169R that can compensate for the lack of other FtsZ bundling factors in vivo.
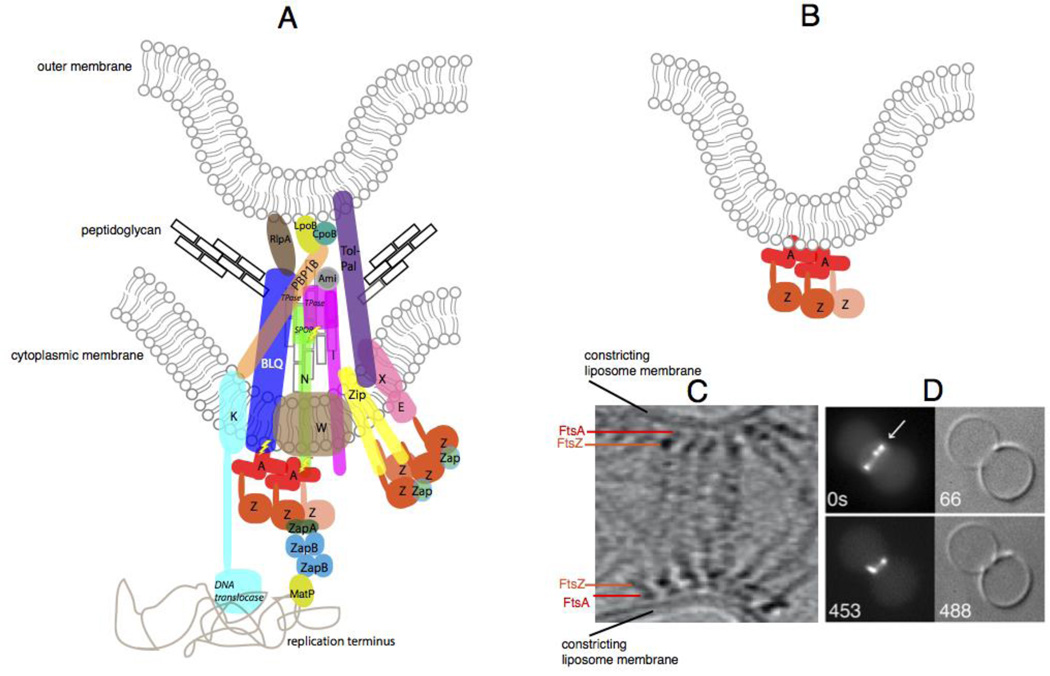
The constricting divisome in vivo and reconstituted in vitro. (A) A schematic of a putative individual sub-complex at the constricting divisome of E. coli shows FtsZ polymers being anchored to the membrane by FtsA and ZipA, along with other essential and nonessential divisome proteins of E. coli that cross the inner membrane, some of which contact the peptidoglycan and the outer membrane. Sidewall and septal peptidoglycan are denoted by black or grey rectangles, respectively. All Fts proteins are shown as a single letter, including the FtsBLQ complex. FtsZ dynamics are shown by the lighter colored FtsZ monomer coming off the polymer. Most known direct protein-protein contacts are shown, but some are speculative. Known enzymatic activities are highlighted in italics (TPase = transpeptidase). ZapC and ZapD are denoted as “Zap”, and the several periplasmic amidases of E. coli are denoted as “Ami”. Also shown are contacts made between the divisome and the terminus region of the nucleoid, including the helicase domain of FtsK and MatP, which probably interacts with ZapB . Regions of putative regulatory signaling are shown with lightning bolts. (B) Schematic of constriction of liposomes in vitro by FtsZ tethered to FtsA in the liposome lumen. (C) Cryo-ET of T. maritima FtsZ and FtsA proteins pinching a liposome from the inside reproduced with permission from Ref . and D) and fluorescence/DIC microscopy of E. coli FtsZ-YFP and FtsA* also pinching liposomes from the inside; reproduced with permission from ref .
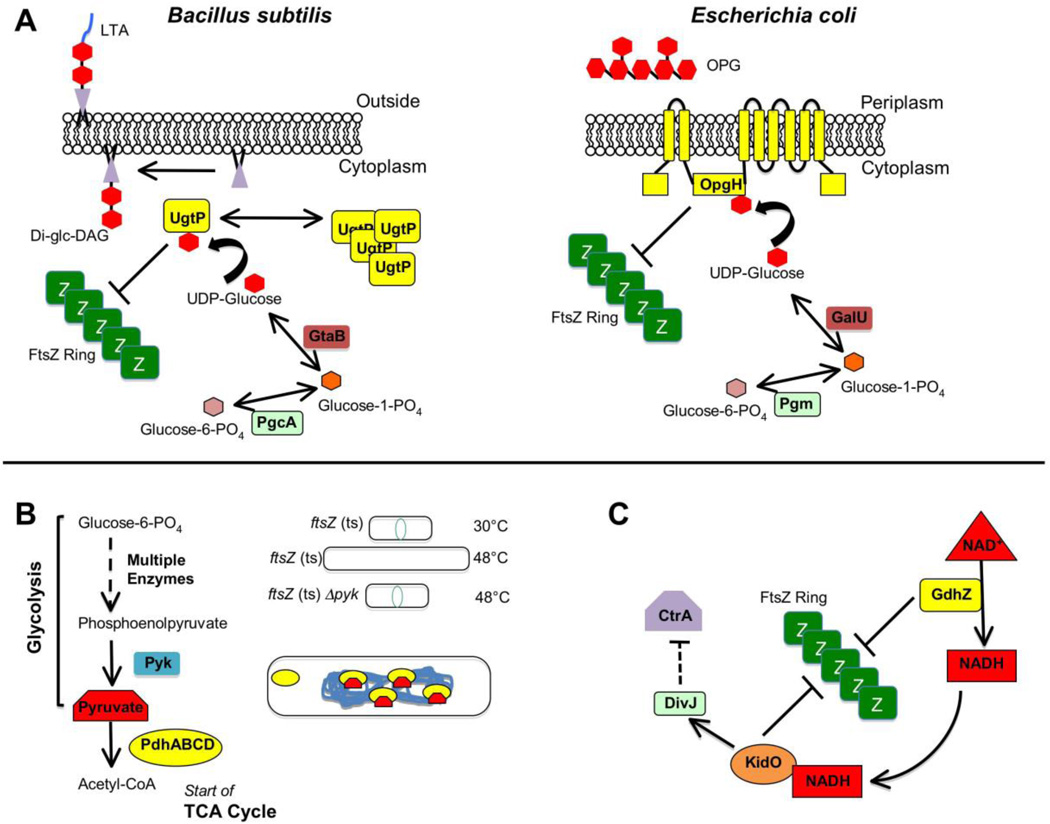
Regulation of divisome activity by stress and nutritional status. (A) The evolutionarily divergent B. subtilis and E. coli each use unrelated glucosyltransferases to coordinate growth rate with cell division through a metabolic determinant: levels of UDP-glucose ,,. For B. subtilis (left), high levels of UDP-glucose during nutrient-rich growth permits UgtP synthesize the diglucosyl-diacylglycerol (Di-glc-DAG) anchor for lipoteichoic acid (LTA), an anionic polymer that is a major component of the Gram-positive cell wall. Additionally some UDP-glucose-bound UgtP interacts with FtsZ and inhibits FtsZ assembly through an uncertain mechanism, delaying division until cells have reached appropriate size for their rapid growth rate. In the absence of UDP-glucose substrate, UgtP is sequestered from FtsZ in cytoplasmic puncta. For E. coli (right), OpgH is a sugar transferase involved in synthesis of osmoregulated periplasmic glucans (OPGs). As with its functional counterpart in Bacillus, OpgH inhibits FtsZ ring formation in a UDP-glucose-dependent manner, likely through sequestration of FtsZ monomers. Although OpgH also only seems to interact with FtsZ during nutrient-rich growth conditions, it does so independently of UDP-glucose, suggesting the existence of other metabolic determinants for this coordination in E. coli. (Modified with permission from ref. ). (B) Enzymes and intermediates involved in glycolysis and the TCA cycle (left) also play a role in B. subtilis cell division . Loss of the pyk gene encoding pyruvate kinase suppresses division defects of a temperature sensitive ftsZ allele under non-permissive conditions (upper & mid, right). In the cell (lower right), binding of pyruvate substrate by the E1alpha subunit (encoded by pdhA) of pyruvate dehydrogenase permits localization of the enzyme/substrate complex to the bacterial nucleoid. E1alpha subunits unable to bind pyruvate substrate (such as in a pyk-null mutant) no longer associate with the nucleoid. (C) A simplification of the highly complex integrated genetic circuit in Caulobacter crescenus linking cell division with metabolism and DNA replication ,. Active GdhZ promotes FtsZ disassembly through stimulation of GTP hydrolysis by FtsZ and through its catalysis of glutamate catabolism into alpha-ketoglutarate provides NADH substrate for KidO, which in turn also can inhibit FtsZ assembly through an unknown mechanism. KidO stimulates the kinase activity of DivJ, which leads to a signal transduction pathway leading through multiple mechanisms to inhibition of the activity of CtrA, a master regulator of both cell division and DNA replication timing in C. crescentus. Activated DivJ additionally feeds back on this circuit to inhibit KidO and GdhZ through proteolytic control governed by ClpXP (not shown).
References
-
- Egan AJ, Vollmer W. The physiology of bacterial cell division. Ann NY Acad Sci. 2013;1277:8–28. - PubMed
-
- Margolin W. Themes and variations in prokaryotic cell division. FEMS Microbiol Rev. 2000;24:531–548. - PubMed
-
- Leisch N, et al. Growth in width and FtsZ ring longitudinal positioning in a gammaproteobacterial symbiont. Curr Biol. 2012;22:R831–R832. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
