

mTORC1 and CK2 coordinate ternary and eIF4F complex assembly
- PMID: 27040916
- PMCID: PMC4822005
- DOI: 10.1038/ncomms11127
mTORC1 and CK2 coordinate ternary and eIF4F complex assembly
Abstract
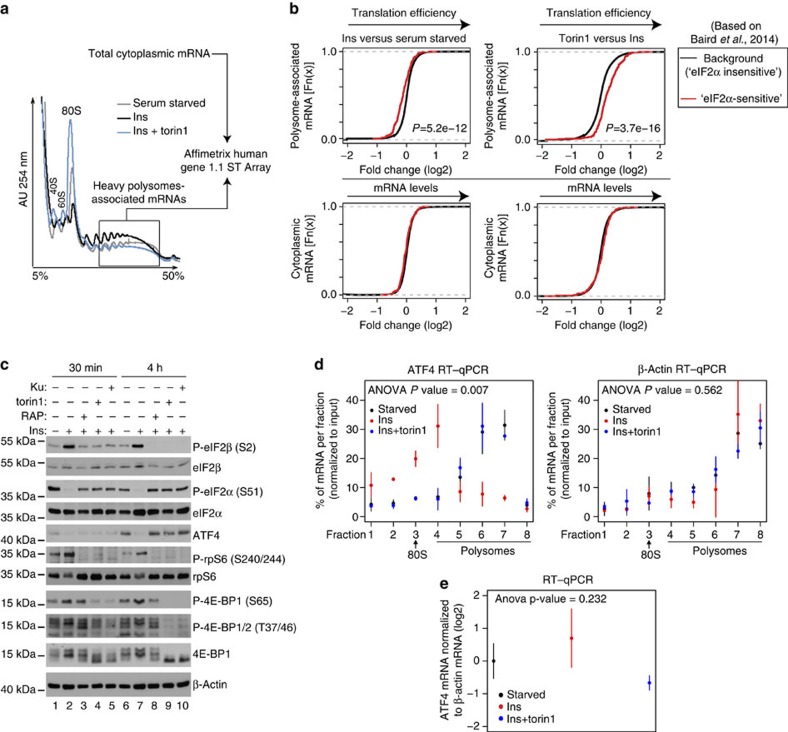
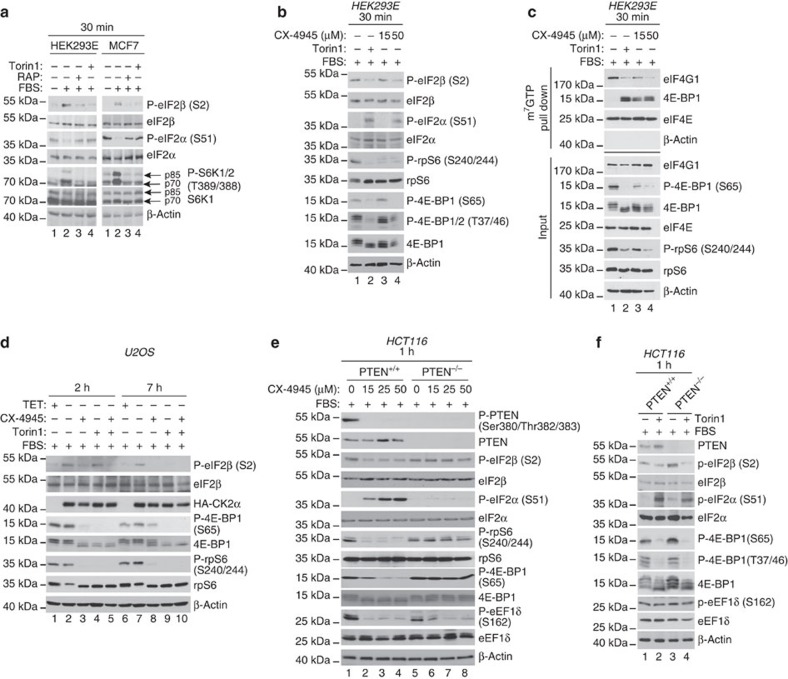
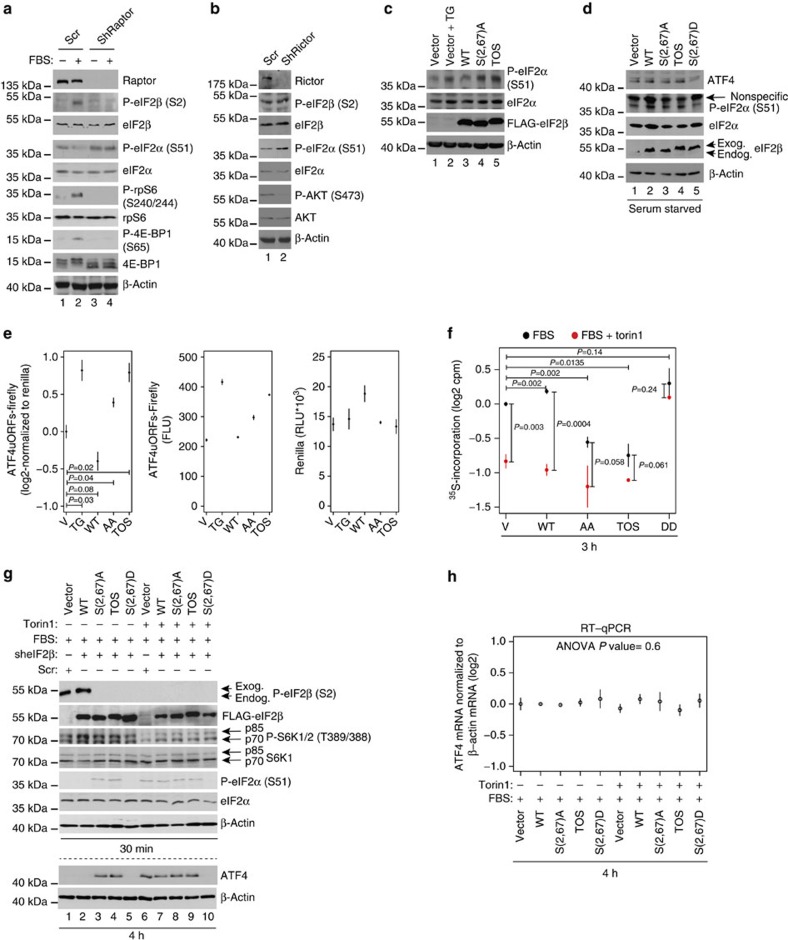
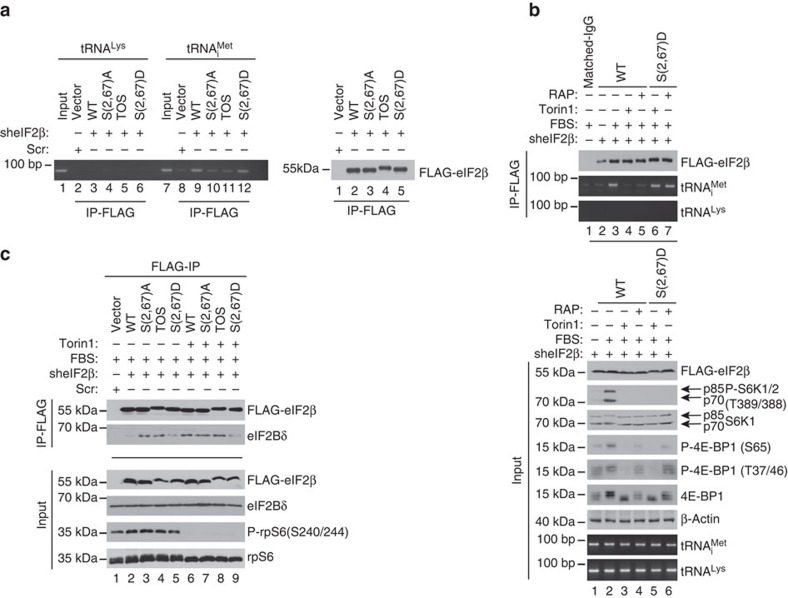
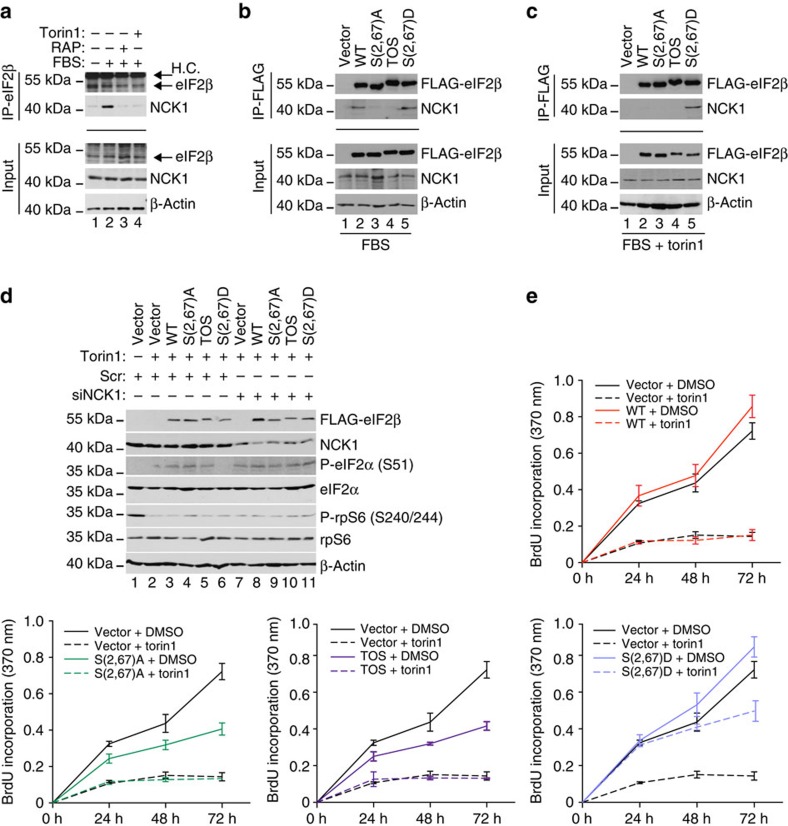
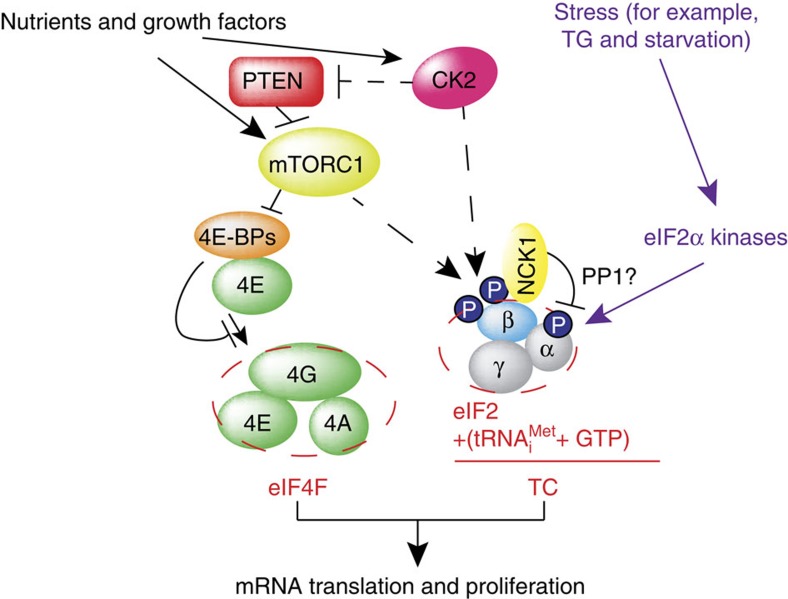
Ternary complex (TC) and eIF4F complex assembly are the two major rate-limiting steps in translation initiation regulated by eIF2α phosphorylation and the mTOR/4E-BP pathway, respectively. How TC and eIF4F assembly are coordinated, however, remains largely unknown. We show that mTOR suppresses translation of mRNAs activated under short-term stress wherein TC recycling is attenuated by eIF2α phosphorylation. During acute nutrient or growth factor stimulation, mTORC1 induces eIF2β phosphorylation and recruitment of NCK1 to eIF2, decreases eIF2α phosphorylation and bolsters TC recycling. Accordingly, eIF2β mediates the effect of mTORC1 on protein synthesis and proliferation. In addition, we demonstrate a formerly undocumented role for CK2 in regulation of translation initiation, whereby CK2 stimulates phosphorylation of eIF2β and simultaneously bolsters eIF4F complex assembly via the mTORC1/4E-BP pathway. These findings imply a previously unrecognized mode of translation regulation, whereby mTORC1 and CK2 coordinate TC and eIF4F complex assembly to stimulate cell proliferation.
Figures
Similar articles
-
Hepatitis C virus NS5A binds to the mRNA cap-binding eukaryotic translation initiation 4F (eIF4F) complex and up-regulates host translation initiation machinery through eIF4E-binding protein 1 inactivation.J Biol Chem. 2012 Feb 10;287(7):5042-58. doi: 10.1074/jbc.M111.308916. Epub 2011 Dec 19. J Biol Chem. 2012. PMID: 22184107 Free PMC article.
-
Sensitivity of global translation to mTOR inhibition in REN cells depends on the equilibrium between eIF4E and 4E-BP1.PLoS One. 2011;6(12):e29136. doi: 10.1371/journal.pone.0029136. Epub 2011 Dec 22. PLoS One. 2011. PMID: 22216185 Free PMC article.
-
Alkaline intracellular pH (pHi) increases PI3K activity to promote mTORC1 and mTORC2 signaling and function during growth factor limitation.J Biol Chem. 2023 Sep;299(9):105097. doi: 10.1016/j.jbc.2023.105097. Epub 2023 Jul 26. J Biol Chem. 2023. PMID: 37507012 Free PMC article.
-
4E-BP1, a multifactor regulated multifunctional protein.Cell Cycle. 2016;15(6):781-6. doi: 10.1080/15384101.2016.1151581. Cell Cycle. 2016. PMID: 26901143 Free PMC article. Review.
-
eIF4F: a retrospective.J Biol Chem. 2015 Oct 2;290(40):24091-9. doi: 10.1074/jbc.R115.675280. Epub 2015 Aug 31. J Biol Chem. 2015. PMID: 26324716 Free PMC article. Review.
Cited by
-
Cap-dependent translation initiation monitored in living cells.Nat Commun. 2022 Nov 2;13(1):6558. doi: 10.1038/s41467-022-34052-8. Nat Commun. 2022. PMID: 36323665 Free PMC article.
-
Surviving and Adapting to Stress: Translational Control and the Integrated Stress Response.Antioxid Redox Signal. 2023 Aug;39(4-6):351-373. doi: 10.1089/ars.2022.0123. Epub 2023 May 9. Antioxid Redox Signal. 2023. PMID: 36943285 Free PMC article. Review.
-
Casein kinase II-dependent phosphorylation of DNA topoisomerase II suppresses the effect of a catalytic topo II inhibitor, ICRF-193, in fission yeast.J Biol Chem. 2019 Mar 8;294(10):3772-3782. doi: 10.1074/jbc.RA118.004955. Epub 2019 Jan 11. J Biol Chem. 2019. PMID: 30635402 Free PMC article.
-
CK2 and the Hallmarks of Cancer.Biomedicines. 2022 Aug 16;10(8):1987. doi: 10.3390/biomedicines10081987. Biomedicines. 2022. PMID: 36009534 Free PMC article. Review.
-
Translation Control by p53.Cancers (Basel). 2018 May 5;10(5):133. doi: 10.3390/cancers10050133. Cancers (Basel). 2018. PMID: 29734756 Free PMC article.
References
-
- Hinnebusch A. G. The scanning mechanism of eukaryotic translation initiation. Annu. Rev. Biochem. 83, 779–812 (2014). - PubMed
-
- Albert V. & Hall M. N. mTOR signaling in cellular and organismal energetics. Curr. Opin. Cell Biol. 33, 55–66 (2015). - PubMed
-
- Koromilas A. E. Roles of the translation initiation factor eIF2alpha serine 51 phosphorylation in cancer formation and treatment. Biochim. Biophys. Acta 1849, 871–880 (2015). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
