

RSK2 signals through stathmin to promote microtubule dynamics and tumor metastasis
- PMID: 27041561
- PMCID: PMC5226067
- DOI: 10.1038/onc.2016.79
RSK2 signals through stathmin to promote microtubule dynamics and tumor metastasis
Abstract
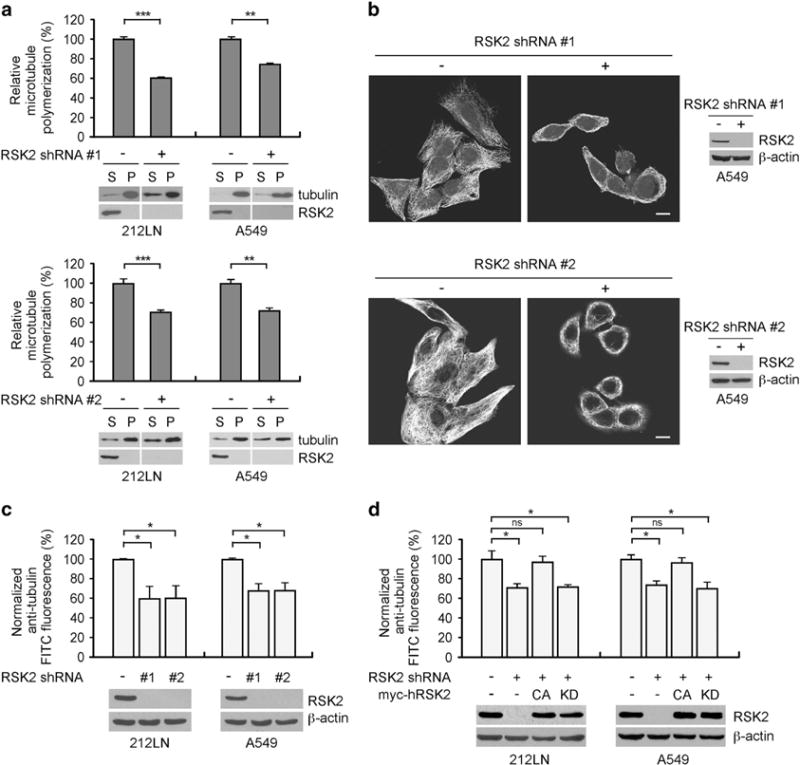
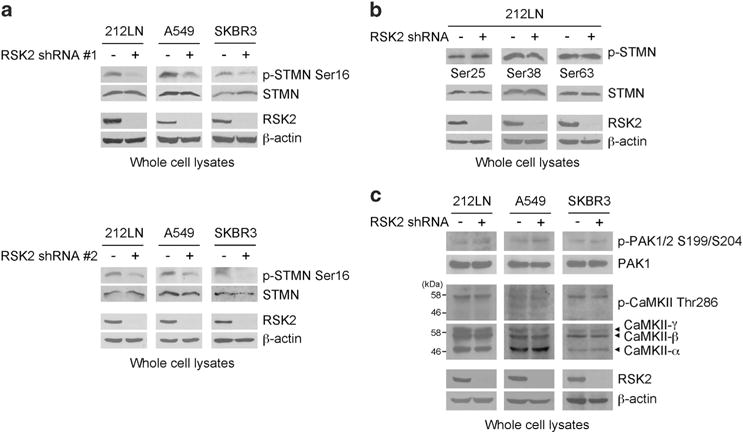
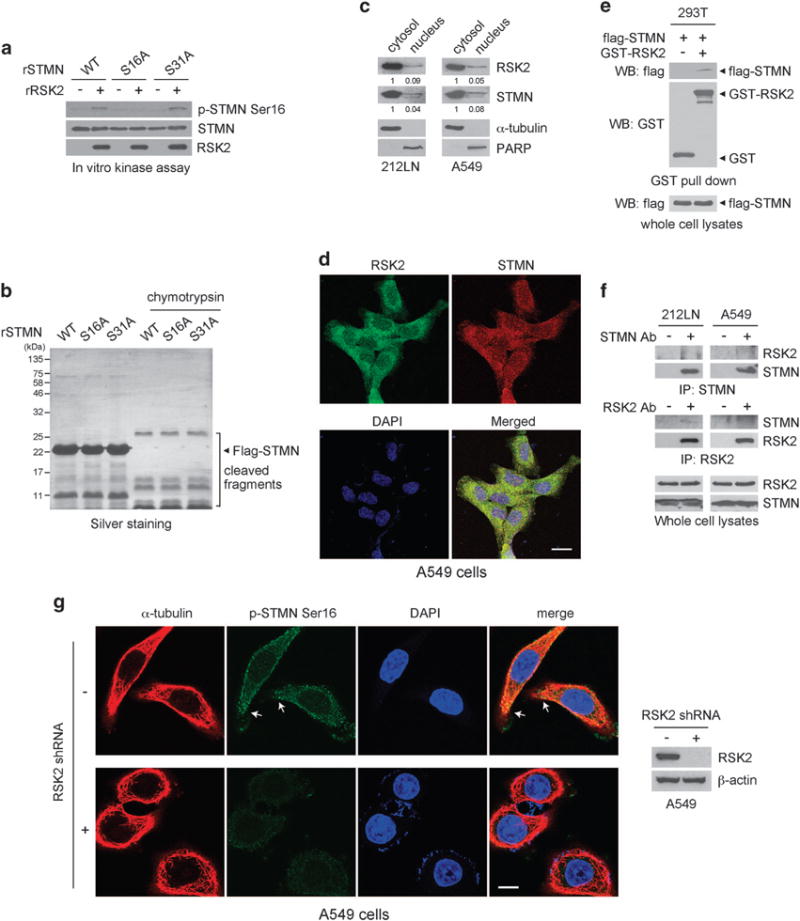
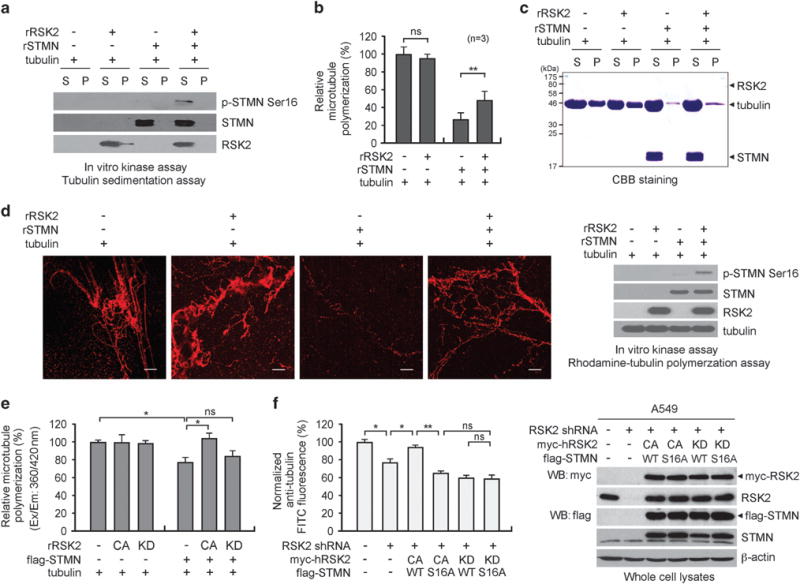
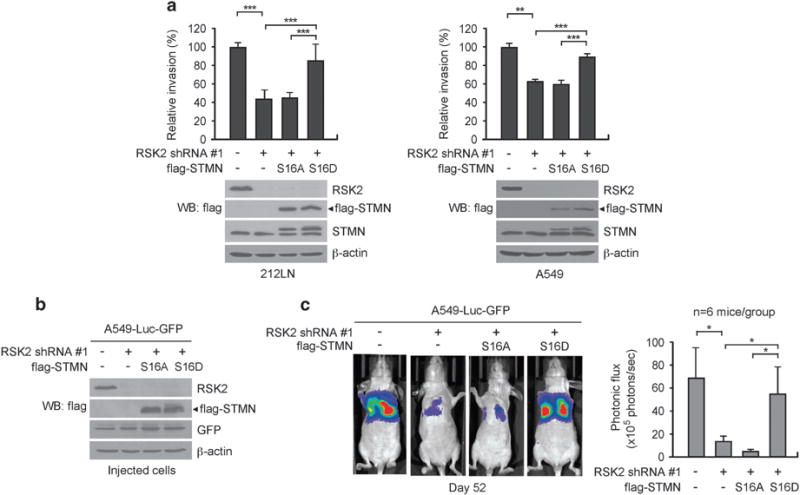
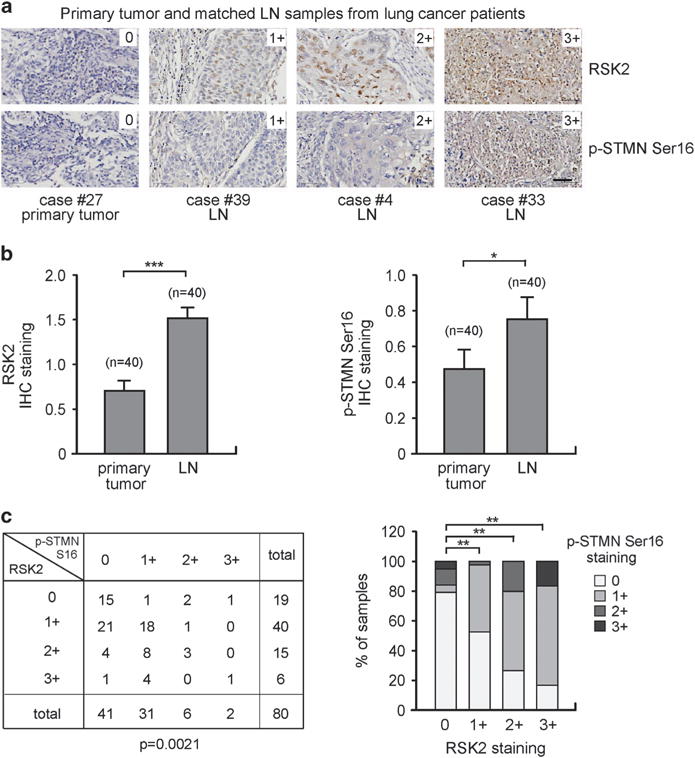
Metastasis is responsible for >90% of cancer-related deaths. Complex signaling in cancer cells orchestrates the progression from a primary to a metastatic cancer. However, the mechanisms of these cellular changes remain elusive. We previously demonstrated that p90 ribosomal S6 kinase 2 (RSK2) promotes tumor metastasis. Here we investigated the role of RSK2 in the regulation of microtubule dynamics and its potential implication in cancer cell invasion and tumor metastasis. Stable knockdown of RSK2 disrupted microtubule stability and decreased phosphorylation of stathmin, a microtubule-destabilizing protein, at serine 16 in metastatic human cancer cells. We found that RSK2 directly binds and phosphorylates stathmin at the leading edge of cancer cells. Phosphorylation of stathmin by RSK2 reduced stathmin-mediated microtubule depolymerization. Moreover, overexpression of phospho-mimetic mutant stathmin S16D significantly rescued the decreased invasive and metastatic potential mediated by RSK2 knockdown in vitro and in vivo. Furthermore, stathmin phosphorylation positively correlated with RSK2 expression and metastatic cancer progression in primary patient tumor samples. Our finding demonstrates that RSK2 directly phosphorylates stathmin and regulates microtubule polymerization to provide a pro-invasive and pro-metastatic advantage to cancer cells. Therefore, the RSK2-stathmin pathway represents a promising therapeutic target and a prognostic marker for metastatic human cancers.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Fidler IJ. The pathogenesis of cancer metastasis: the ‘seed and soil’ hypothesis revisited. Nat Rev Cancer. 2003;3:453–458. - PubMed
-
- Nguyen DX, Massague J. Genetic determinants of cancer metastasis. Nat Rev Genet. 2007;8:341–352. - PubMed
-
- Gupta GP, Massague J. Cancer metastasis: building a framework. Cell. 2006;127:679–695. - PubMed
-
- Sahai E. Mechanisms of cancer cell invasion. Curr Opin Genet Dev. 2005;15:87–96. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
