

Transcriptome Remodeling in Trypanosoma cruzi and Human Cells during Intracellular Infection
- PMID: 27046031
- PMCID: PMC4821583
- DOI: 10.1371/journal.ppat.1005511
Transcriptome Remodeling in Trypanosoma cruzi and Human Cells during Intracellular Infection
Abstract
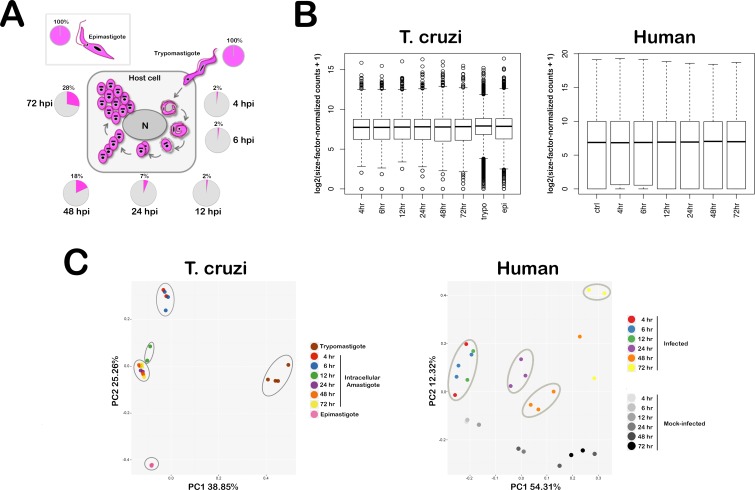
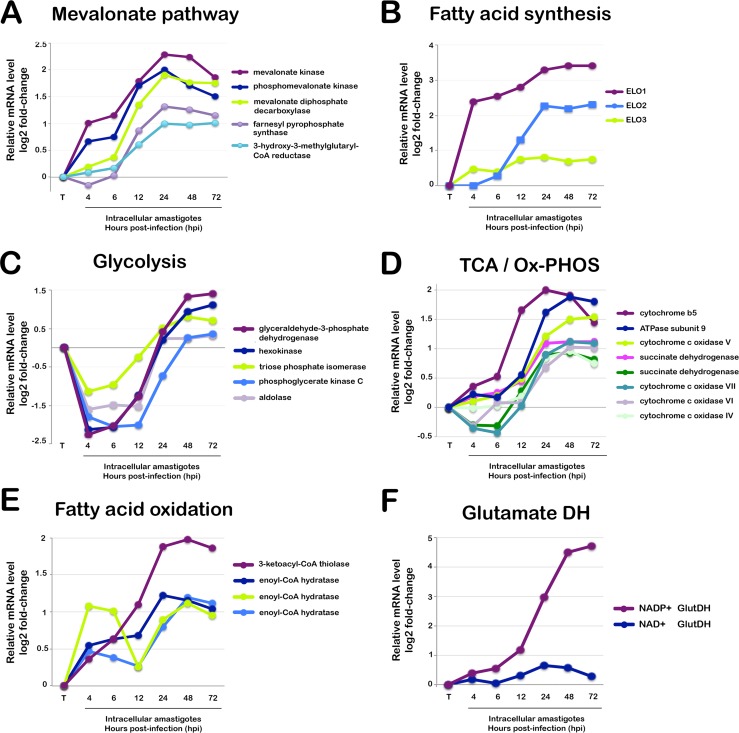
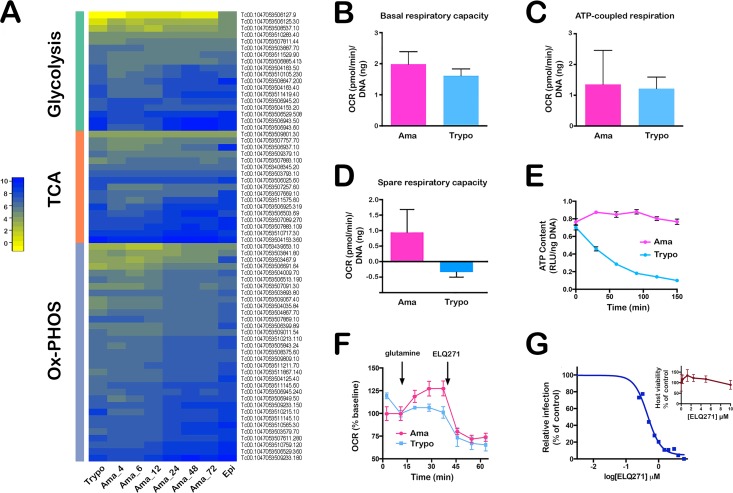
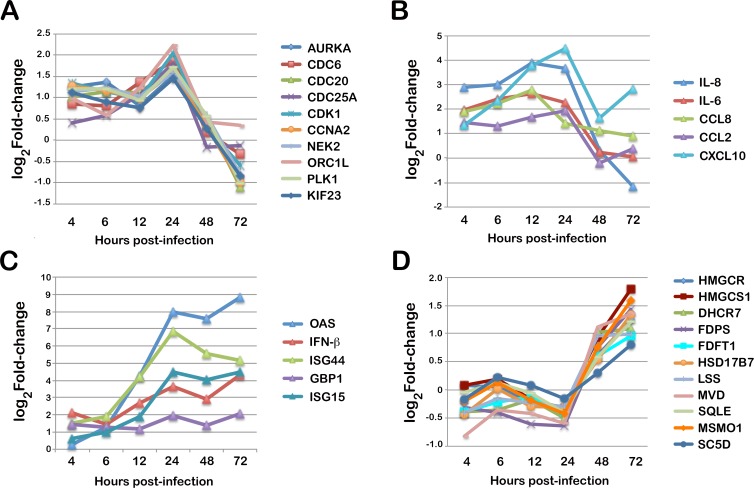
Intracellular colonization and persistent infection by the kinetoplastid protozoan parasite, Trypanosoma cruzi, underlie the pathogenesis of human Chagas disease. To obtain global insights into the T. cruzi infective process, transcriptome dynamics were simultaneously captured in the parasite and host cells in an infection time course of human fibroblasts. Extensive remodeling of the T. cruzi transcriptome was observed during the early establishment of intracellular infection, coincident with a major developmental transition in the parasite. Contrasting this early response, few additional changes in steady state mRNA levels were detected once mature T. cruzi amastigotes were formed. Our findings suggest that transcriptome remodeling is required to establish a modified template to guide developmental transitions in the parasite, whereas homeostatic functions are regulated independently of transcriptomic changes, similar to that reported in related trypanosomatids. Despite complex mechanisms for regulation of phenotypic expression in T. cruzi, transcriptomic signatures derived from distinct developmental stages mirror known or projected characteristics of T. cruzi biology. Focusing on energy metabolism, we were able to validate predictions forecast in the mRNA expression profiles. We demonstrate measurable differences in the bioenergetic properties of the different mammalian-infective stages of T. cruzi and present additional findings that underscore the importance of mitochondrial electron transport in T. cruzi amastigote growth and survival. Consequences of T. cruzi colonization for the host include dynamic expression of immune response genes and cell cycle regulators with upregulation of host cholesterol and lipid synthesis pathways, which may serve to fuel intracellular T. cruzi growth. Thus, in addition to the biological inferences gained from gene ontology and functional enrichment analysis of differentially expressed genes in parasite and host, our comprehensive, high resolution transcriptomic dataset provides a substantially more detailed interpretation of T. cruzi infection biology and offers a basis for future drug and vaccine discovery efforts.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Tardieux I, Webster P, Ravesloot J, Boron W, Lunn JA, Heuser JE, et al. Lysosome recruitment and fusion are early events required for trypanosome invasion of mammalian cells. Cell. 1992;71(7):1117–30. - PubMed
-
- Tomlinson S, Vandekerckhove F, Frevert U, Nussenzweig V. The induction of Trypanosoma cruzi trypomastigote to amastigote transformation by low pH. Parasitology. 1995;110 (Pt 5):547–54. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
