

Septin oligomerization regulates persistent expression of ErbB2/HER2 in gastric cancer cells
- PMID: 27048593
- PMCID: PMC4903893
- DOI: 10.1042/BCJ20160203
Septin oligomerization regulates persistent expression of ErbB2/HER2 in gastric cancer cells
Abstract
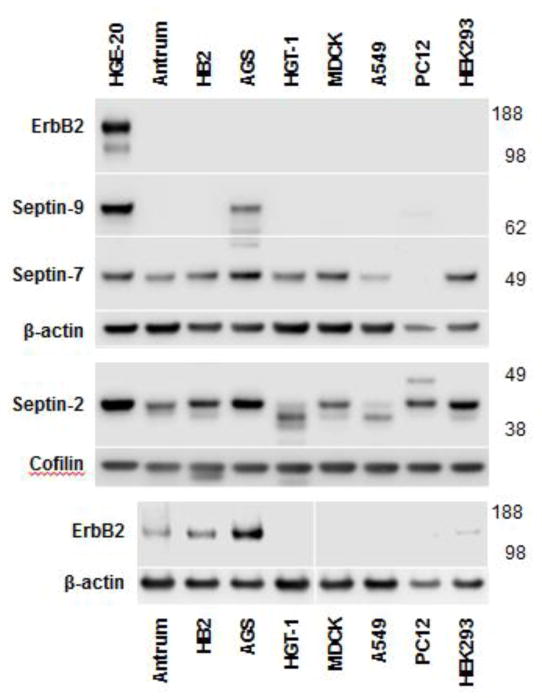
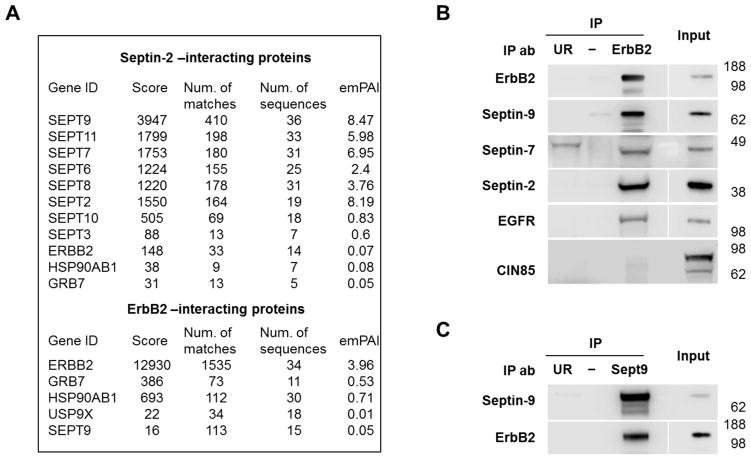
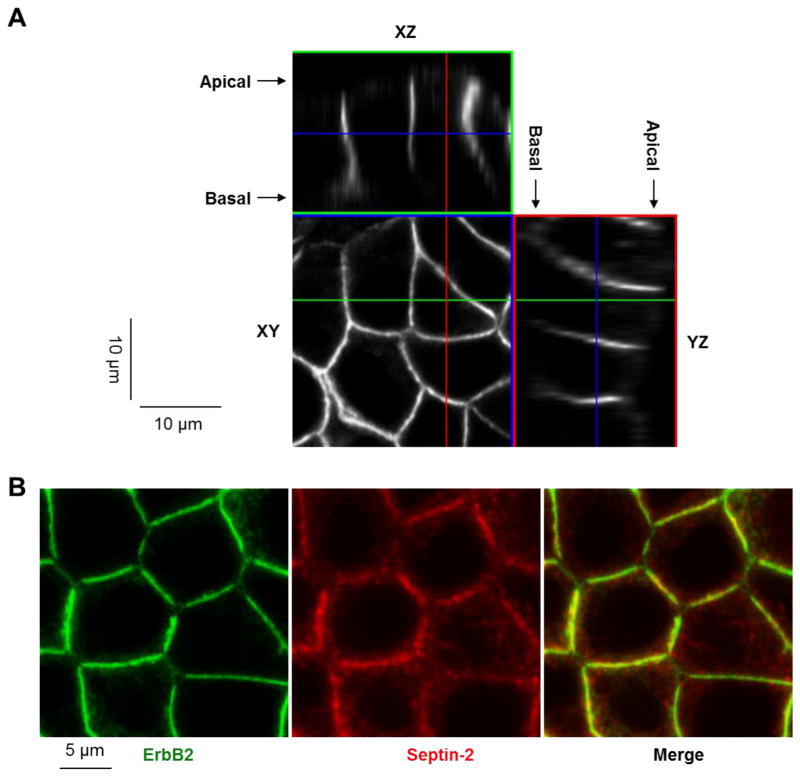
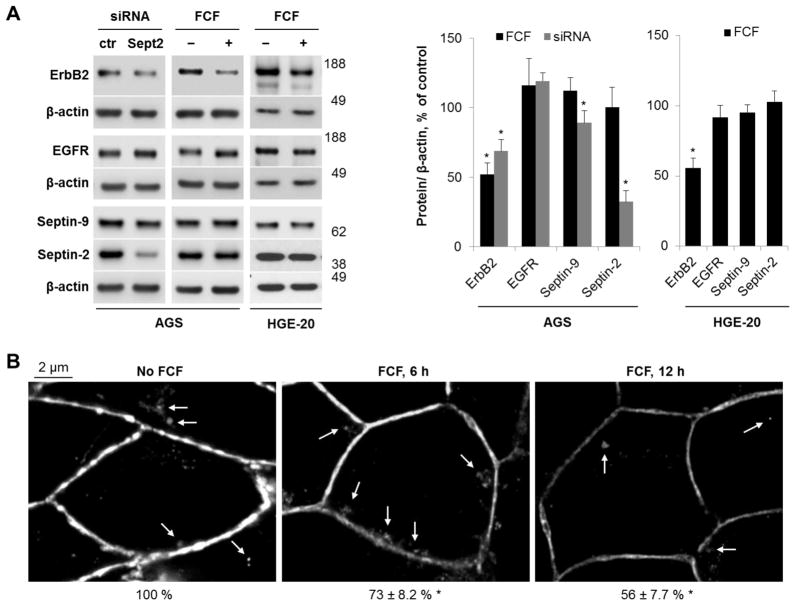
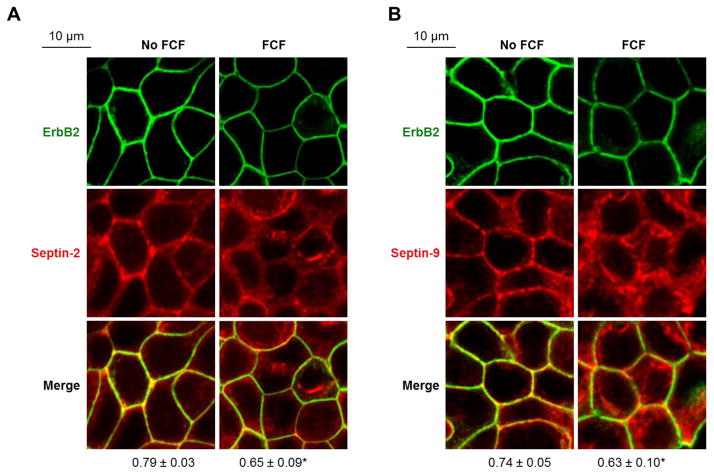
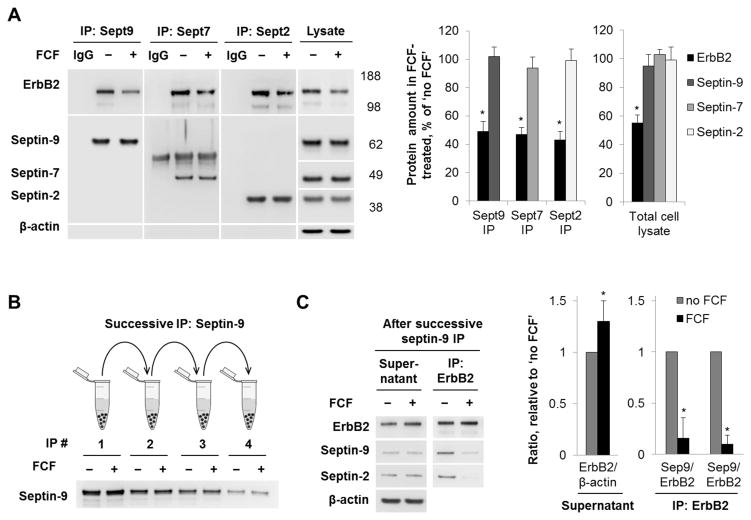
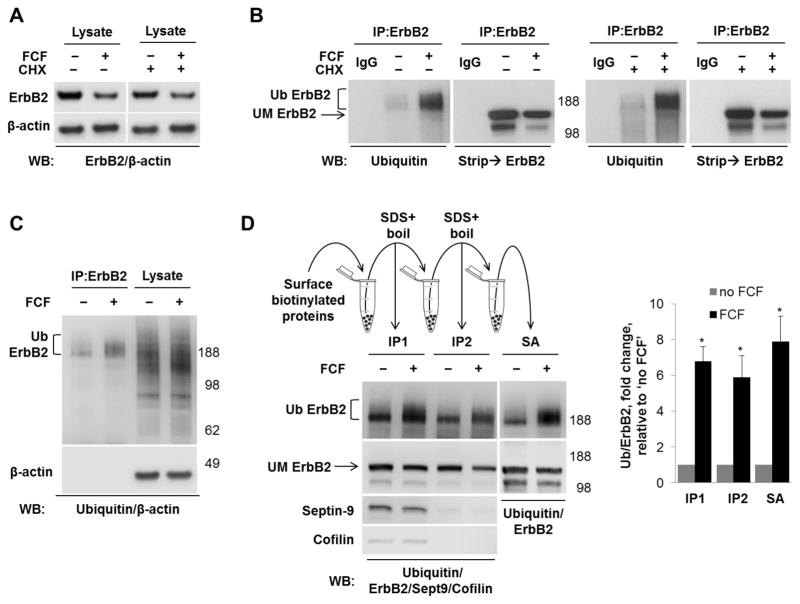
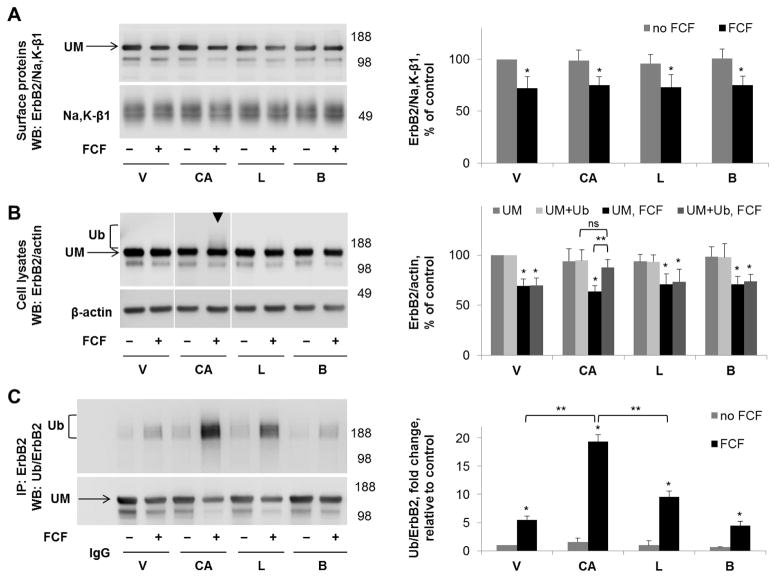
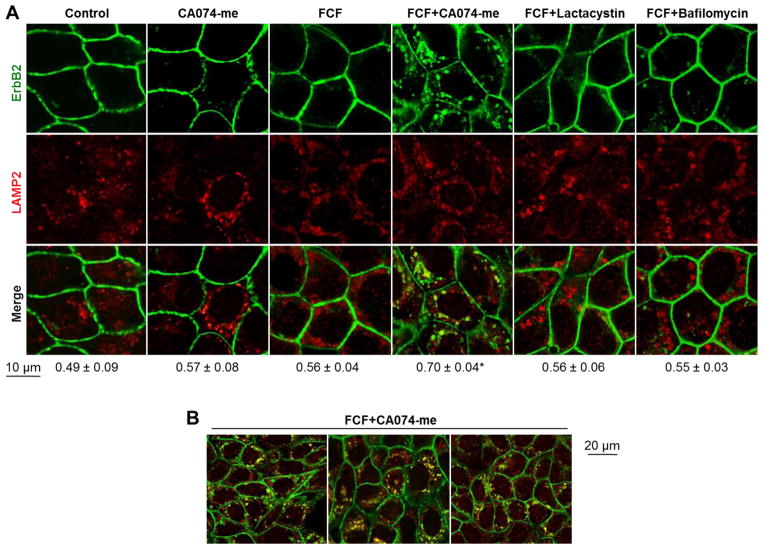
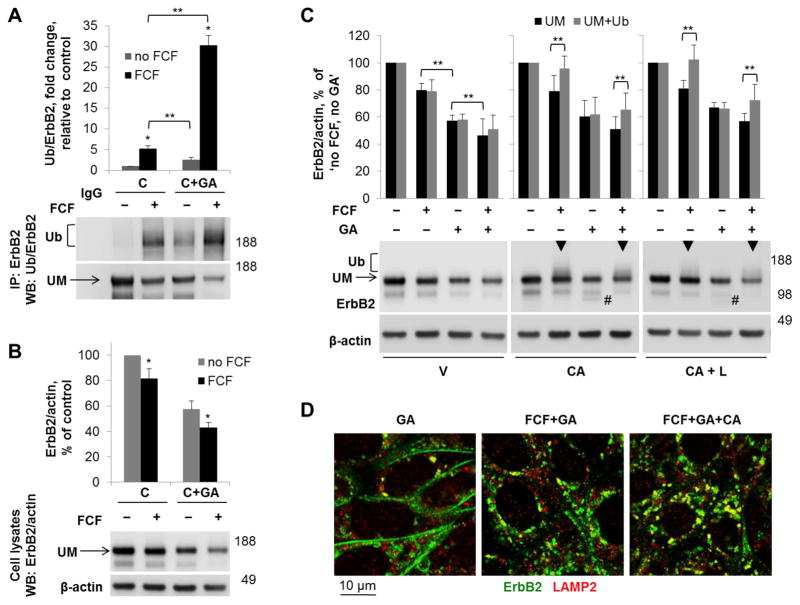
Septins are a family of cytoskeletal GTP-binding proteins that assemble into membrane-associated hetero-oligomers and organize scaffolds for recruitment of cytosolic proteins or stabilization of membrane proteins. Septins have been implicated in a diverse range of cancers, including gastric cancer, but the underlying mechanisms remain unclear. The hypothesis tested here is that septins contribute to cancer by stabilizing the receptor tyrosine kinase ErbB2, an important target for cancer treatment. Septins and ErbB2 were highly over-expressed in gastric cancer cells. Immunoprecipitation followed by MS analysis identified ErbB2 as a septin-interacting protein. Knockdown of septin-2 or cell exposure to forchlorfenuron (FCF), a well-established inhibitor of septin oligomerization, decreased surface and total levels of ErbB2. These treatments had no effect on epidermal growth factor receptor (EGFR), emphasizing the specificity and functionality of the septin-ErbB2 interaction. The level of ubiquitylated ErbB2 at the plasma membrane was elevated in cells treated with FCF, which was accompanied by a decrease in co-localization of ErbB2 with septins at the membrane. Cathepsin B inhibitor, but not bafilomycin or lactacystin, prevented FCF-induced decrease in total ErbB2 by increasing accumulation of ubiquitylated ErbB2 in lysosomes. Therefore, septins protect ErbB2 from ubiquitylation, endocytosis and lysosomal degradation. The FCF-induced degradation pathway is distinct from and additive with the degradation induced by inhibiting ErbB2 chaperone Hsp90. These results identify septins as novel regulators of ErbB2 expression that contribute to the remarkable stabilization of the receptor at the plasma membrane of cancer cells and may provide a basis for the development of new ErbB2-targeting anti-cancer therapies.
Keywords: ErbB2; endocytosis; gastric cancer; lysosomal degradation; septin; ubiquitylation.
© 2016 The Author(s). published by Portland Press Limited on behalf of the Biochemical Society.
Conflict of interest statement
Figures
Similar articles
-
A Septin Cytoskeleton-Targeting Small Molecule, Forchlorfenuron, Inhibits Epithelial Migration via Septin-Independent Perturbation of Cellular Signaling.Cells. 2019 Dec 29;9(1):84. doi: 10.3390/cells9010084. Cells. 2019. PMID: 31905721 Free PMC article.
-
Septin dynamics are essential for exocytosis.J Biol Chem. 2015 Feb 27;290(9):5280-97. doi: 10.1074/jbc.M114.616201. Epub 2015 Jan 9. J Biol Chem. 2015. PMID: 25575596 Free PMC article.
-
Recruitment of septin cytoskeletal proteins by botulinum toxin A protease determines its remarkable stability.J Cell Sci. 2014 Aug 1;127(Pt 15):3294-308. doi: 10.1242/jcs.146324. Epub 2014 Jun 13. J Cell Sci. 2014. PMID: 24928902 Free PMC article.
-
[Functional Characterization of Septin Complexes].Mol Biol (Mosk). 2018 Mar-Apr;52(2):155-171. doi: 10.7868/S0026898418020015. Mol Biol (Mosk). 2018. PMID: 29695686 Free PMC article. Review.
-
Septins in animal tissue architecture: more than just peanuts.Curr Opin Cell Biol. 2025 Jun;94:102525. doi: 10.1016/j.ceb.2025.102525. Epub 2025 Apr 30. Curr Opin Cell Biol. 2025. PMID: 40311264 Review.
Cited by
-
Septin-2 is overexpressed in epithelial ovarian cancer and mediates proliferation via regulation of cellular metabolic proteins.Oncotarget. 2019 Apr 26;10(31):2959-2972. doi: 10.18632/oncotarget.26836. eCollection 2019 Apr 26. Oncotarget. 2019. PMID: 31105878 Free PMC article.
-
Septins: Regulators of Protein Stability.Front Cell Dev Biol. 2016 Dec 20;4:143. doi: 10.3389/fcell.2016.00143. eCollection 2016. Front Cell Dev Biol. 2016. PMID: 28066764 Free PMC article. Review.
-
Development of Potent Forchlorfenuron Analogs and Their Cytotoxic Effect in Cancer Cell Lines.Sci Rep. 2020 Feb 24;10(1):3241. doi: 10.1038/s41598-020-59824-4. Sci Rep. 2020. PMID: 32094384 Free PMC article.
-
Cancer-Related Functions and Subcellular Localizations of Septins.Front Cell Dev Biol. 2016 Nov 8;4:126. doi: 10.3389/fcell.2016.00126. eCollection 2016. Front Cell Dev Biol. 2016. PMID: 27878118 Free PMC article. Review.
-
Septins As Modulators of Endo-Lysosomal Membrane Traffic.Front Cell Dev Biol. 2016 Nov 3;4:124. doi: 10.3389/fcell.2016.00124. eCollection 2016. Front Cell Dev Biol. 2016. PMID: 27857942 Free PMC article. Review.
References
-
- Hall PA, Russell SE. Mammalian septins: dynamic heteromers with roles in cellular morphogenesis and compartmentalization. The Journal of pathology. 2012;226:287–299. - PubMed
-
- Connolly D, Abdesselam I, Verdier-Pinard P, Montagna C. Septin roles in tumorigenesis. Biol Chem. 2011;392:725–738. - PubMed
-
- Scott M, Hyland PL, McGregor G, Hillan KJ, Russell SE, Hall PA. Multimodality expression profiling shows SEPT9 to be overexpressed in a wide range of human tumours. Oncogene. 2005;24:4688–4700. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous