

Resveratrol Attenuates Trimethylamine-N-Oxide (TMAO)-Induced Atherosclerosis by Regulating TMAO Synthesis and Bile Acid Metabolism via Remodeling of the Gut Microbiota
- PMID: 27048804
- PMCID: PMC4817264
- DOI: 10.1128/mBio.02210-15
Resveratrol Attenuates Trimethylamine-N-Oxide (TMAO)-Induced Atherosclerosis by Regulating TMAO Synthesis and Bile Acid Metabolism via Remodeling of the Gut Microbiota
Abstract
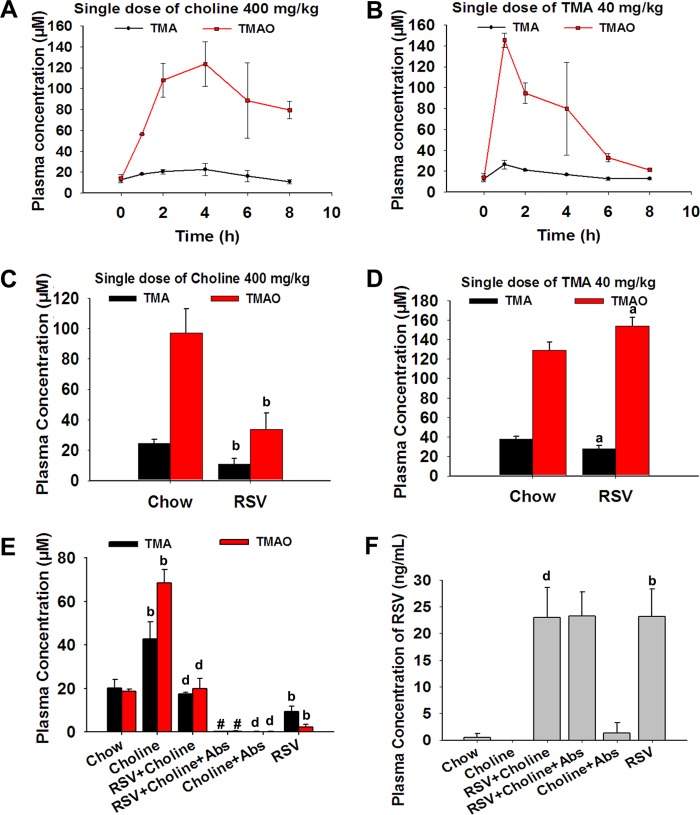
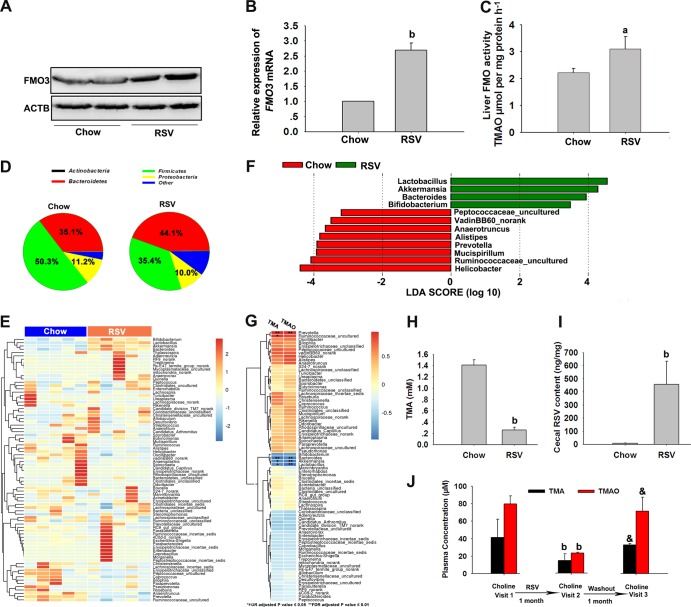
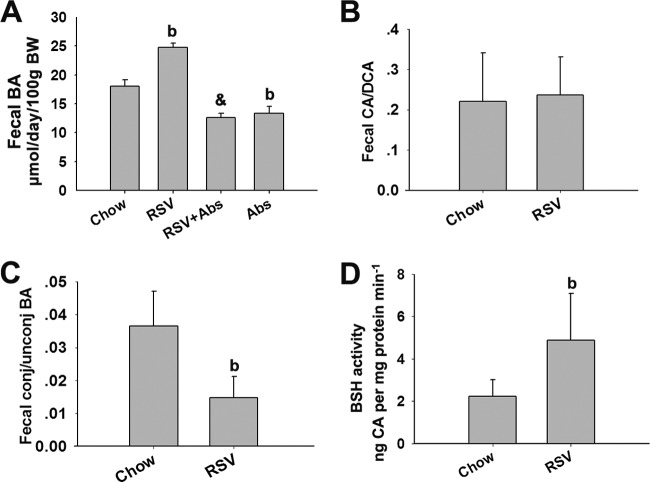
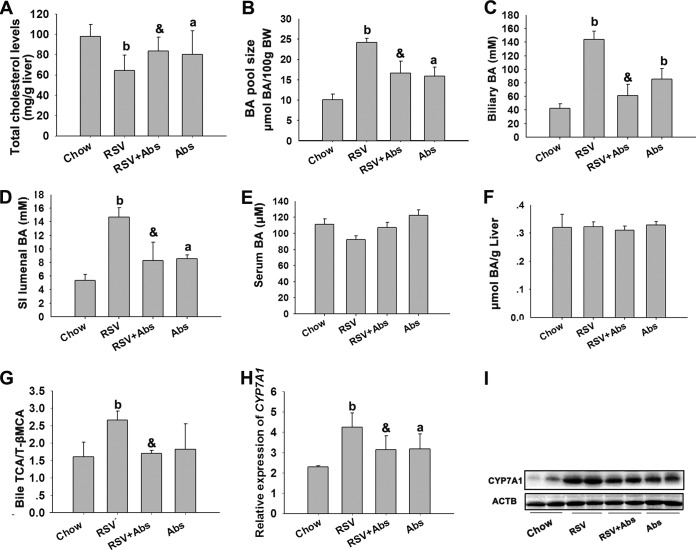
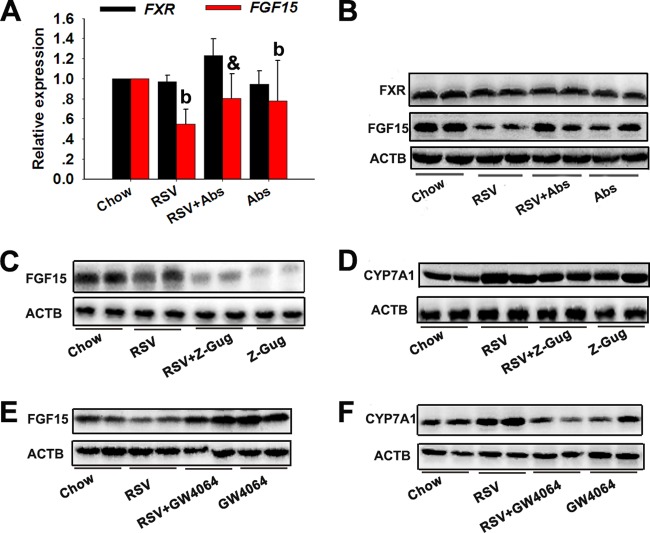
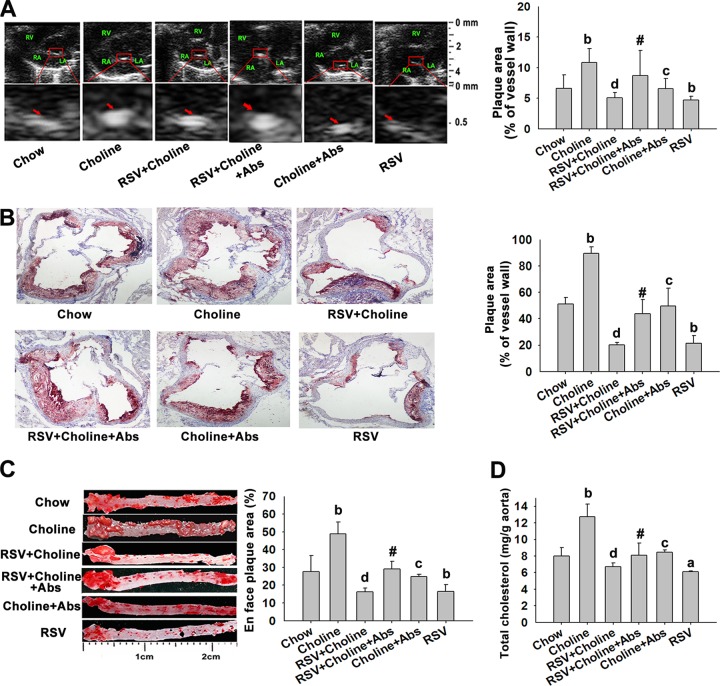
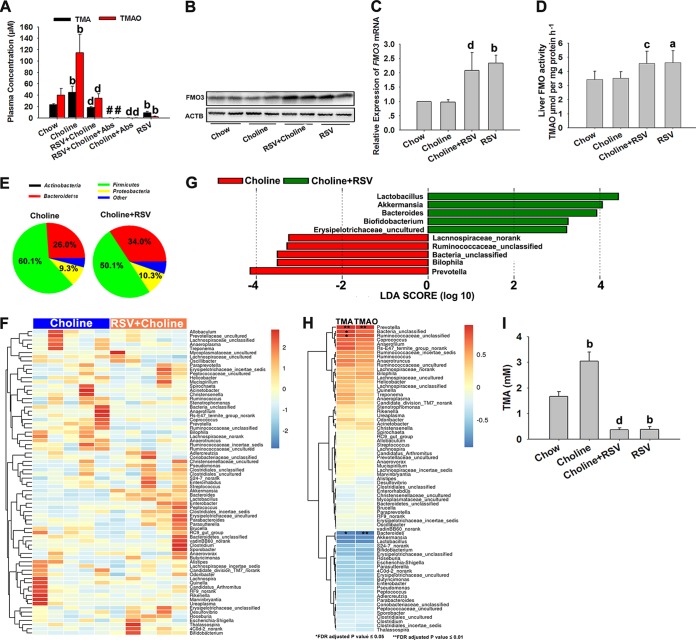
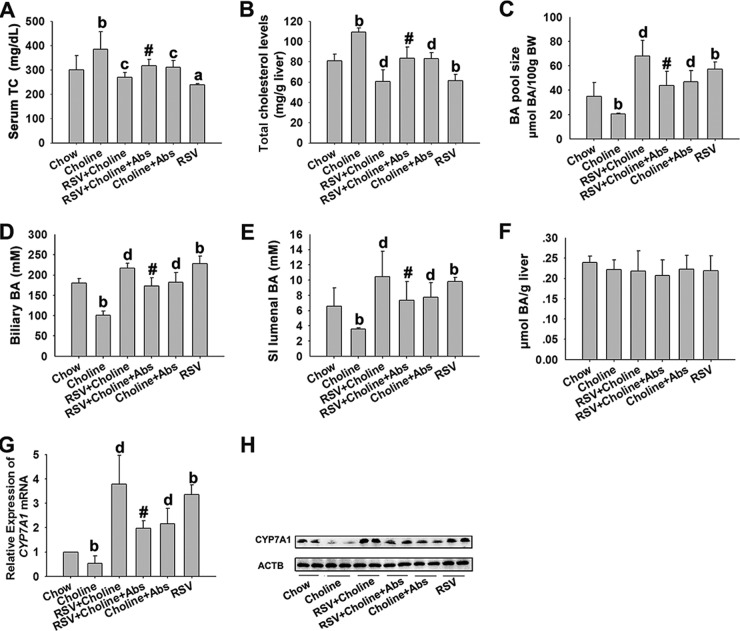
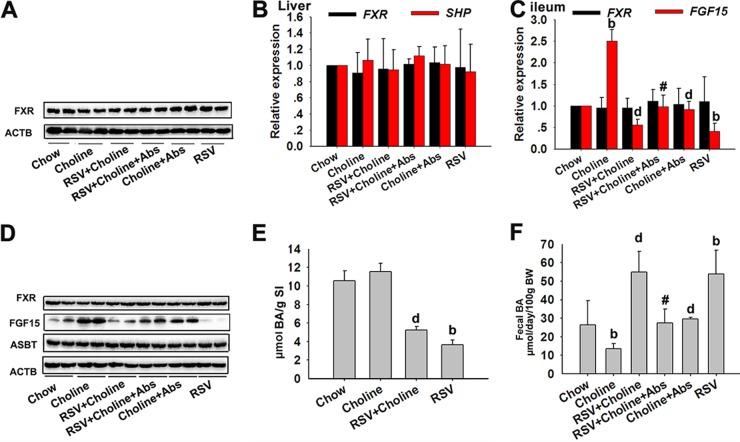
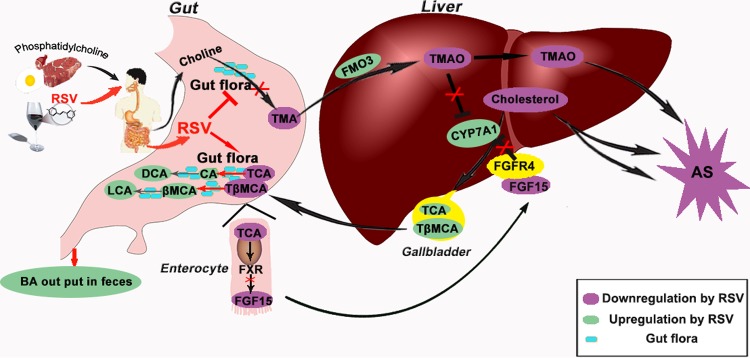
The gut microbiota is found to be strongly associated with atherosclerosis (AS). Resveratrol (RSV) is a natural phytoalexin with anti-AS effects; however, its mechanisms of action remain unclear. Therefore, we sought to determine whether the anti-AS effects of RSV were related to changes in the gut microbiota. We found that RSV attenuated trimethylamine-N-oxide (TMAO)-induced AS in ApoE(-/-) mice. Meanwhile, RSV decreased TMAO levels by inhibiting commensal microbial trimethylamine (TMA) production via gut microbiota remodeling in mice. Moreover, RSV increased levels of the genera Lactobacillus and Bifidobacterium, which increased the bile salt hydrolase activity, thereby enhancing bile acid (BA) deconjugation and fecal excretion in C57BL/6J and ApoE(-/-) mice. This was associated with a decrease in ileal BA content, repression of the enterohepatic farnesoid X receptor (FXR)-fibroblast growth factor 15 (FGF15) axis, and increased cholesterol 7a-hydroxylase (CYP7A1) expression and hepatic BA neosynthesis. An FXR antagonist had the same effect on FGF15 and CYP7A1 expression as RSV, while an FXR agonist abolished RSV-induced alterations in FGF15 and CYP7A1 expression. In mice treated with antibiotics, RSV neither decreased TMAO levels nor increased hepatic BA synthesis. Additionally, RSV-induced inhibition of TMAO-caused AS was also markedly abolished by antibiotics. In conclusion, RSV attenuated TMAO-induced AS by decreasing TMAO levels and increasing hepatic BA neosynthesis via gut microbiota remodeling, and the BA neosynthesis was partially mediated through the enterohepatic FXR-FGF15 axis.
Importance: Recently, trimethylamine-N-oxide (TMAO) has been identified as a novel and independent risk factor for promoting atherosclerosis (AS) partially through inhibiting hepatic bile acid (BA) synthesis. The gut microbiota plays a key role in the pathophysiology of TMAO-induced AS. Resveratrol (RSV) is a natural phytoalexin with prebiotic benefits. A growing body of evidence supports the hypothesis that phenolic phytochemicals with poor bioavailability are possibly acting primarily through remodeling of the gut microbiota. The current study showed that RSV attenuated TMAO-induced AS by decreasing TMAO levels and increasing hepatic BA neosynthesis via gut microbiota remodeling. And RSV-induced hepatic BA neosynthesis was partially mediated through downregulating the enterohepatic farnesoid X receptor-fibroblast growth factor 15 axis. These results offer new insights into the mechanisms responsible for RSV's anti-AS effects and indicate that the gut microbiota may become an interesting target for pharmacological or dietary interventions to decrease the risk of developing cardiovascular diseases.
Copyright © 2016 Chen et al.
Figures
Similar articles
-
Trimethylamine N-Oxide Aggravates Liver Steatosis through Modulation of Bile Acid Metabolism and Inhibition of Farnesoid X Receptor Signaling in Nonalcoholic Fatty Liver Disease.Mol Nutr Food Res. 2019 Sep;63(17):e1900257. doi: 10.1002/mnfr.201900257. Epub 2019 May 22. Mol Nutr Food Res. 2019. PMID: 31095863
-
Ligustrum robustum Alleviates Atherosclerosis by Decreasing Serum TMAO, Modulating Gut Microbiota, and Decreasing Bile Acid and Cholesterol Absorption in Mice.Mol Nutr Food Res. 2021 Jul;65(14):e2100014. doi: 10.1002/mnfr.202100014. Epub 2021 Jun 10. Mol Nutr Food Res. 2021. PMID: 34005835
-
Microbiota modification with probiotics induces hepatic bile acid synthesis via downregulation of the Fxr-Fgf15 axis in mice.Cell Rep. 2014 Apr 10;7(1):12-8. doi: 10.1016/j.celrep.2014.02.032. Epub 2014 Mar 20. Cell Rep. 2014. PMID: 24656817
-
Trimethylamine-N-oxide: a link between the gut microbiome, bile acid metabolism, and atherosclerosis.Curr Opin Lipidol. 2016 Apr;27(2):148-54. doi: 10.1097/MOL.0000000000000274. Curr Opin Lipidol. 2016. PMID: 26959704 Review.
-
Gut microbiota in atherosclerosis: focus on trimethylamine N-oxide.APMIS. 2020 May;128(5):353-366. doi: 10.1111/apm.13038. Epub 2020 Mar 30. APMIS. 2020. PMID: 32108960 Free PMC article. Review.
Cited by
-
Reduction of TMAO level enhances the stability of carotid atherosclerotic plaque through promoting macrophage M2 polarization and efferocytosis.Biosci Rep. 2021 May 28;41(6):BSR20204250. doi: 10.1042/BSR20204250. Biosci Rep. 2021. PMID: 33969376 Free PMC article.
-
FMO3 deficiency of duck leads to decreased lipid deposition and increased antibacterial activity.J Anim Sci Biotechnol. 2022 Nov 16;13(1):119. doi: 10.1186/s40104-022-00777-1. J Anim Sci Biotechnol. 2022. PMID: 36380386 Free PMC article.
-
Implication of Gut Microbiota in Cardiovascular Diseases.Oxid Med Cell Longev. 2020 Sep 26;2020:5394096. doi: 10.1155/2020/5394096. eCollection 2020. Oxid Med Cell Longev. 2020. PMID: 33062141 Free PMC article. Review.
-
Gut Microbiota and Environment in Coronary Artery Disease.Int J Environ Res Public Health. 2021 Apr 16;18(8):4242. doi: 10.3390/ijerph18084242. Int J Environ Res Public Health. 2021. PMID: 33923612 Free PMC article. Review.
-
Role of traditional Chinese medicine in age-related macular degeneration: exploring the gut microbiota's influence.Front Pharmacol. 2024 Jan 25;15:1356324. doi: 10.3389/fphar.2024.1356324. eCollection 2024. Front Pharmacol. 2024. PMID: 38333011 Free PMC article. Review.
References
-
- Wang Z, Klipfell E, Bennett BJ, Koeth R, Levison BS, Dugar B, Feldstein AE, Britt EB, Fu X, Chung YM, Wu Y, Schauer P, Smith JD, Allayee H, Tang WH, DiDonato JA, Lusis AJ, Hazen SL. 2011. Gut flora metabolism of phosphatidylcholine promotes cardiovascular disease. Nature 472:57–63. doi: 10.1038/nature09922. - DOI - PMC - PubMed
-
- Koeth RA, Wang Z, Levison BS, Buffa JA, Org E, Sheehy BT, Britt EB, Fu X, Wu Y, Li L, Smith JD, DiDonato JA, Chen J, Li H, Wu GD, Lewis JD, Warrier M, Brown JM, Krauss RM, Tang WH. 2013. Intestinal microbiota metabolism of l-carnitine, a nutrient in red meat, promotes atherosclerosis. Nat Med 19:576–585. doi: 10.1038/nm.3145. - DOI - PMC - PubMed
-
- Sayin SI, Wahlström A, Felin J, Jäntti S, Marschall HU, Bamberg K, Angelin B, Hyötyläinen T, Orešič M, Bäckhed F. 2013. Gut microbiota regulates bile acid metabolism by reducing the levels of tauro-beta-muricholic acid, a naturally occurring FXR antagonist. Cell Metab 17:225–235. doi: 10.1016/j.cmet.2013.01.003. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
