

Specific Elimination of Latently HIV-1 Infected Cells Using HIV-1 Protease-Sensitive Toxin Nanocapsules
- PMID: 27049645
- PMCID: PMC4822841
- DOI: 10.1371/journal.pone.0151572
Specific Elimination of Latently HIV-1 Infected Cells Using HIV-1 Protease-Sensitive Toxin Nanocapsules
Abstract
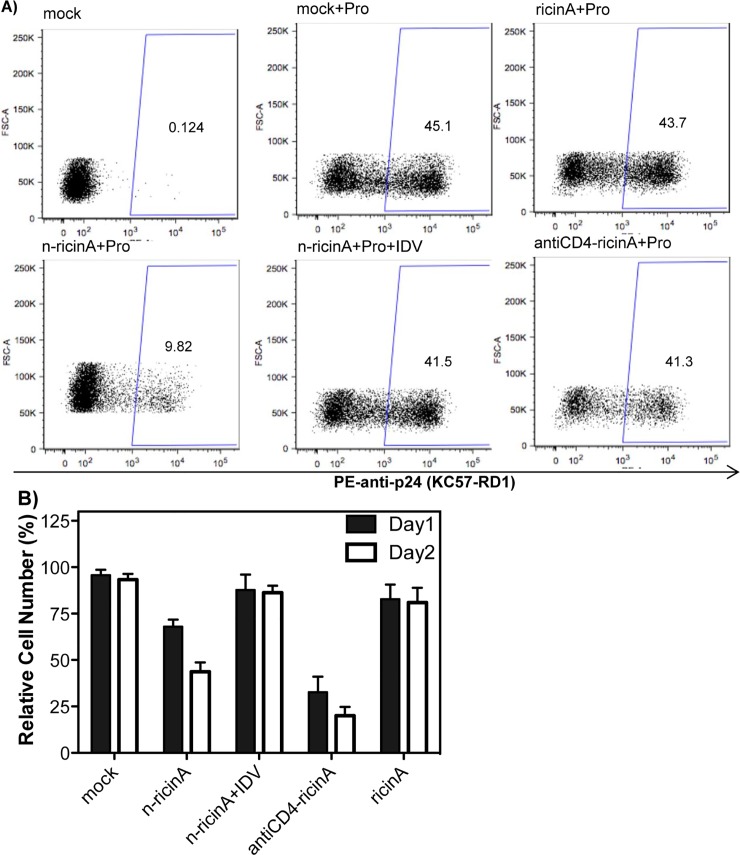
Anti-retroviral drugs suppress HIV-1 plasma viremia to undetectable levels; however, latent HIV-1 persists in reservoirs within HIV-1-infected patients. The silent provirus can be activated through the use of drugs, including protein kinase C activators and histone deacetylase inhibitors. This "shock" approach is then followed by "kill" of the producing cells either through direct HIV-1-induced cell death or natural immune mechanisms. However, these mechanisms are relatively slow and effectiveness is unclear. Here, we develop an approach to specifically target and kill cells that are activated early in the process of virus production. We utilize a novel nanocapsule technology whereby the ricin A chain is encapsulated in an inactive form within a polymer shell. Specificity for release of the ricin A toxin is conferred by peptide crosslinkers that are sensitive to cleavage by HIV-1 protease. By using well-established latent infection models, J-Lat and U1 cells, we demonstrate that only within an HIV-1-producing cell expressing functional HIV-1 protease will the nanocapsule release its ricin A cargo, shutting down viral and cellular protein synthesis, and ultimately leading to rapid death of the producer cell. Thus, we provide proof of principle for a novel technology to kill HIV-1-producing cells without effects on non-target cells.
Conflict of interest statement
Figures






Similar articles
-
Synergistic activation of HIV-1 expression by deacetylase inhibitors and prostratin: implications for treatment of latent infection.PLoS One. 2009 Jun 30;4(6):e6093. doi: 10.1371/journal.pone.0006093. PLoS One. 2009. PMID: 19564922 Free PMC article.
-
Reservoirs for HIV-1: mechanisms for viral persistence in the presence of antiviral immune responses and antiretroviral therapy.Annu Rev Immunol. 2000;18:665-708. doi: 10.1146/annurev.immunol.18.1.665. Annu Rev Immunol. 2000. PMID: 10837072 Review.
-
Ingenol derivates promising for HIV eradication.AIDS Rev. 2014 Oct-Dec;16(4):246. AIDS Rev. 2014. PMID: 25373349
-
In vivo analysis of the effect of panobinostat on cell-associated HIV RNA and DNA levels and latent HIV infection.Retrovirology. 2016 May 21;13(1):36. doi: 10.1186/s12977-016-0268-7. Retrovirology. 2016. PMID: 27206407 Free PMC article.
-
The development of immune-modulating compounds to disrupt HIV latency.Cytokine Growth Factor Rev. 2012 Aug-Oct;23(4-5):159-72. doi: 10.1016/j.cytogfr.2012.05.003. Epub 2012 Jul 4. Cytokine Growth Factor Rev. 2012. PMID: 22766356 Review.
Cited by
-
Plant Ribosome-Inactivating Proteins: Progesses, Challenges and Biotechnological Applications (and a Few Digressions).Toxins (Basel). 2017 Oct 12;9(10):314. doi: 10.3390/toxins9100314. Toxins (Basel). 2017. PMID: 29023422 Free PMC article. Review.
-
Pan-retroviral Nucleocapsid-Mediated Phase Separation Regulates Genomic RNA Positioning and Trafficking.Cell Rep. 2020 Apr 21;31(3):107520. doi: 10.1016/j.celrep.2020.03.084. Cell Rep. 2020. PMID: 32320662 Free PMC article.
-
Impact of Chronic HIV/SIV Infection on T Follicular Helper Cell Subsets and Germinal Center Homeostasis.Front Immunol. 2016 Nov 11;7:501. doi: 10.3389/fimmu.2016.00501. eCollection 2016. Front Immunol. 2016. PMID: 27891132 Free PMC article. Review.
-
U1 and OM10.1. Myeloid Cell Lines as Surrogate Models of Reversible Proviral Latency.Methods Mol Biol. 2022;2407:17-28. doi: 10.1007/978-1-0716-1871-4_2. Methods Mol Biol. 2022. PMID: 34985654
-
Targeted Nanocarrier Delivery of RNA Therapeutics to Control HIV Infection.Pharmaceutics. 2022 Jun 26;14(7):1352. doi: 10.3390/pharmaceutics14071352. Pharmaceutics. 2022. PMID: 35890248 Free PMC article. Review.
References
-
- Gulick RM, Mellors JW, Havlir D, Eron JJ, Gonzalez C, McMahon D, et al. Treatment with indinavir, zidovudine, and lamivudine in adults with human immunodeficiency virus infection and prior antiretroviral therapy. New Engl J Med. 1997;337(11):734–739. - PubMed
-
- Perelson AS, Essunger P, Cao YZ, Vesanen M, Hurley A, Saksela K, et al. Decay characteristics of HIV-1-infected compartments during combination therapy. Nature. 1997;387(6629):188–191. - PubMed
-
- Finzi D, Blankson J, Siliciano JD, Margolick JB, Chadwick K, Pierson T, et al. Latent infection of CD4(+) T cells provides a mechanism for lifelong persistence of HIV-1, even in patients on effective combination therapy. Nature Medicine. 1999;5(5):512–517. - PubMed
-
- Ruff CT, Ray SC, Kwon P, Zinn R, Pendleton A, Hutton N, et al. Persistence of Wild-Type Virus and Lack of Temporal Structure in the Latent Reservoir for Human Immunodeficiency Virus Type 1 in Pediatric Patients with Extensive Antiretroviral Exposure. Journal of Virology. 2002;76(18):9481–9492. - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
