

2D and 3D Stem Cell Models of Primate Cortical Development Identify Species-Specific Differences in Progenitor Behavior Contributing to Brain Size
- PMID: 27049876
- PMCID: PMC4826446
- DOI: 10.1016/j.stem.2016.03.003
2D and 3D Stem Cell Models of Primate Cortical Development Identify Species-Specific Differences in Progenitor Behavior Contributing to Brain Size
Abstract
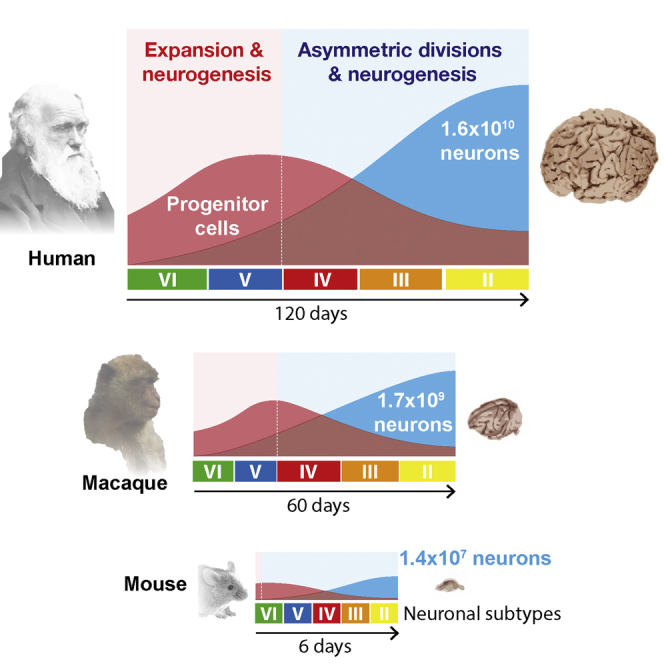
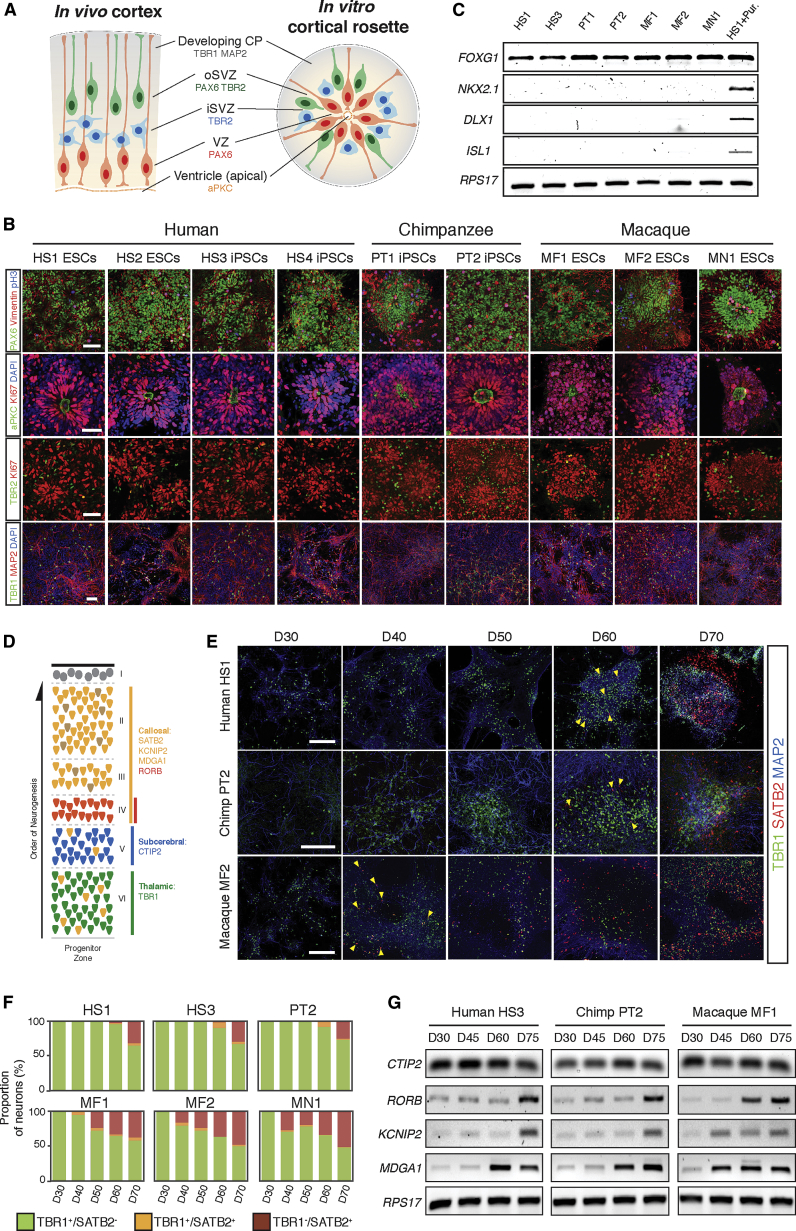
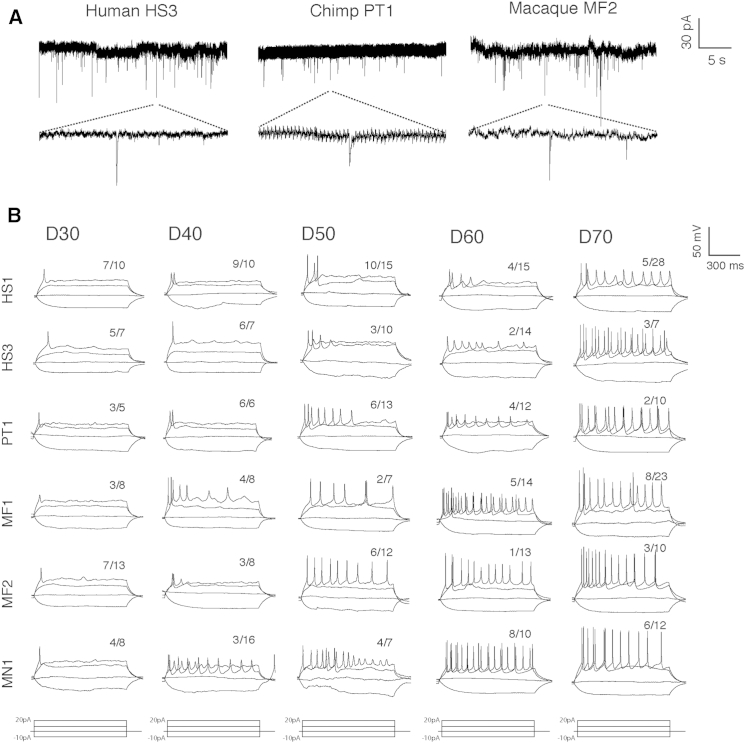
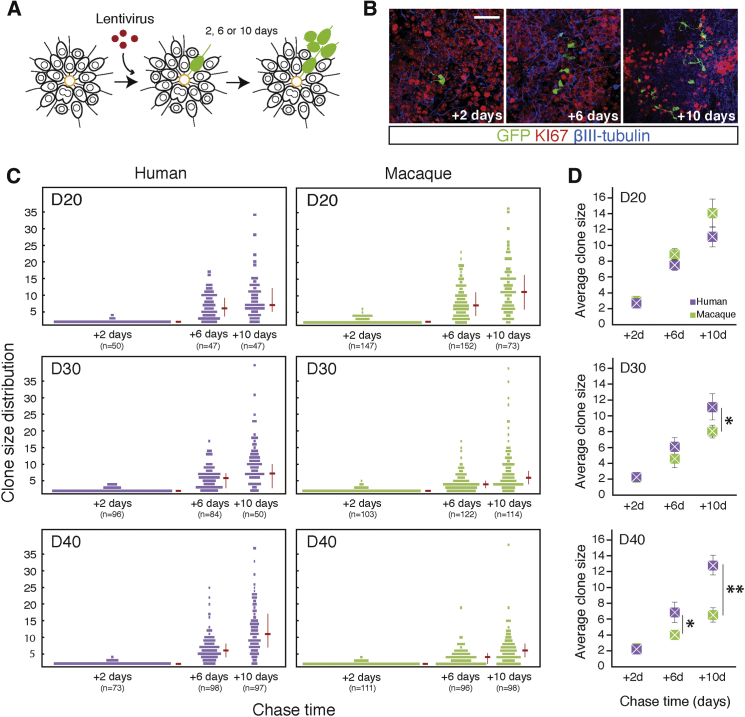
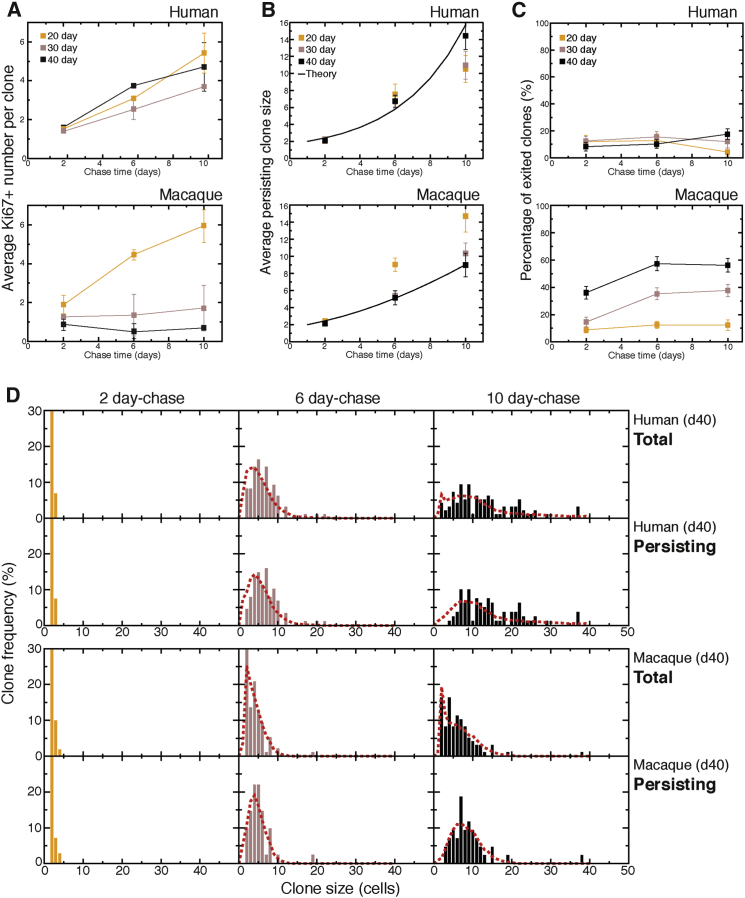
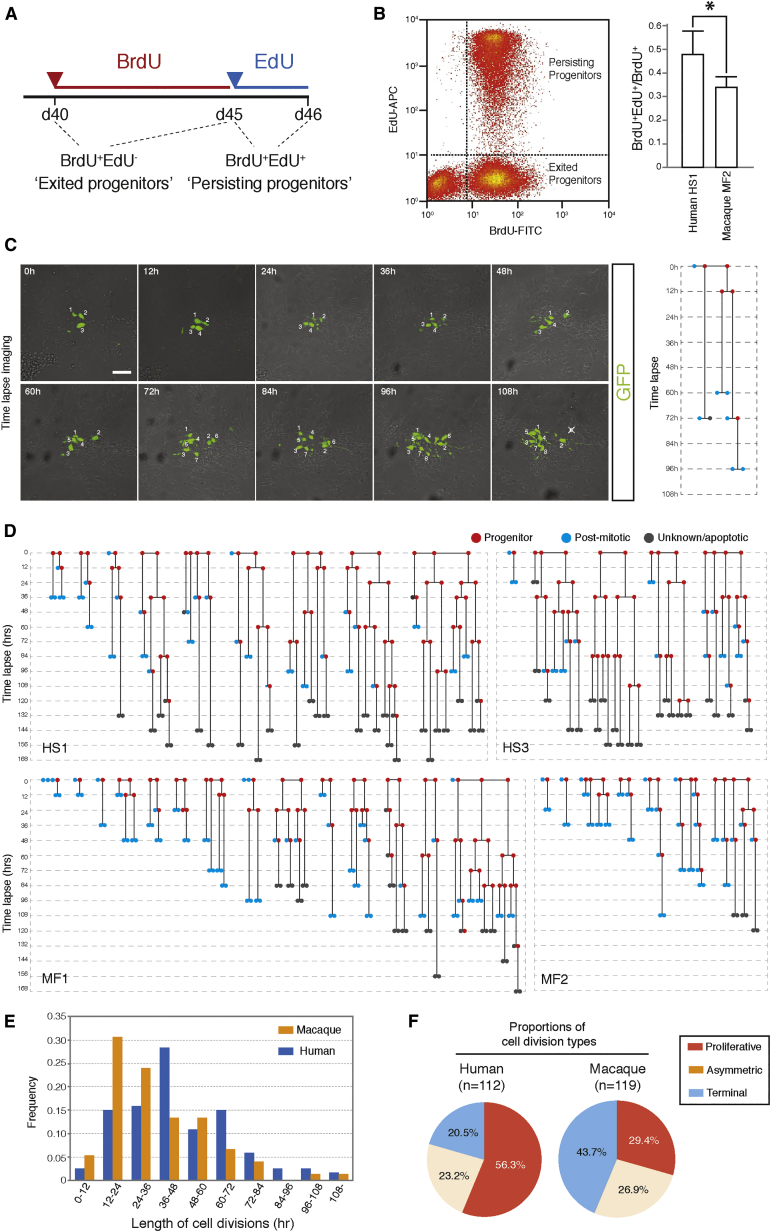
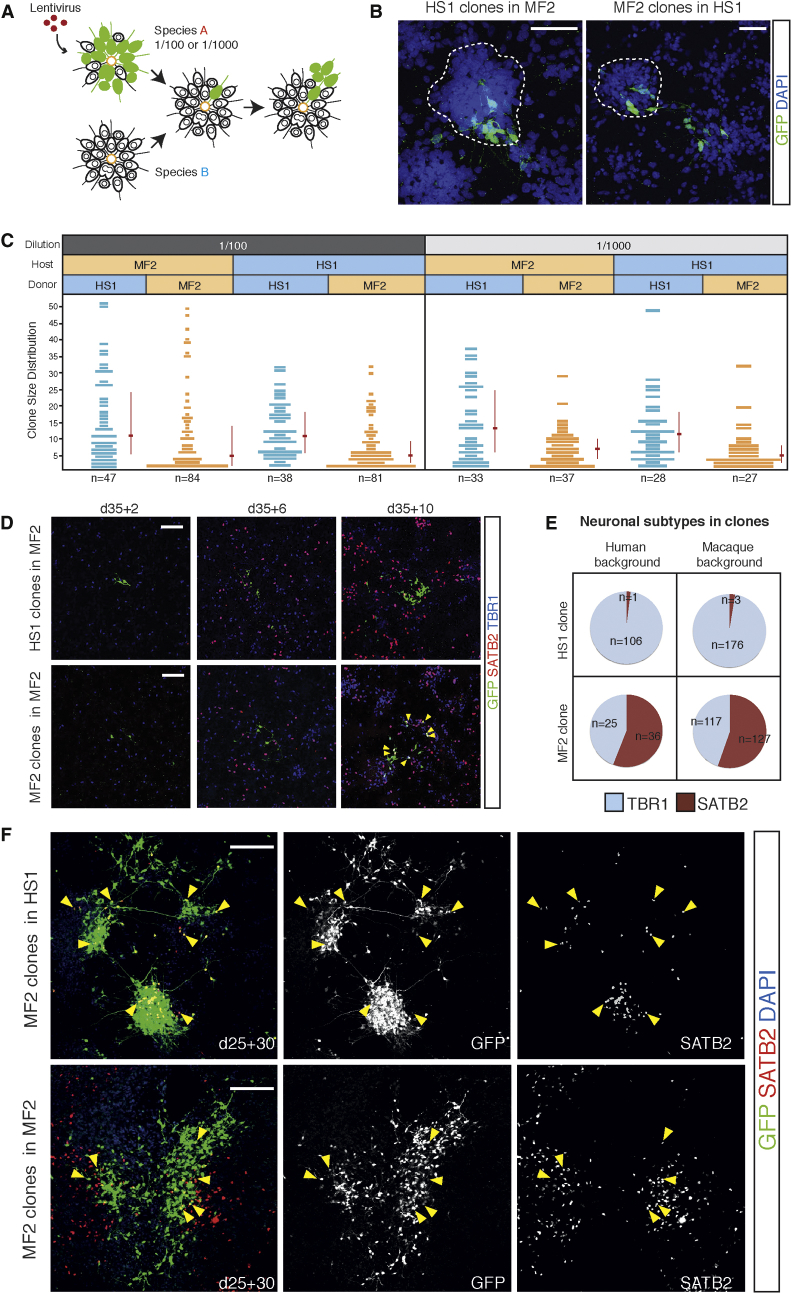
Variation in cerebral cortex size and complexity is thought to contribute to differences in cognitive ability between humans and other animals. Here we compare cortical progenitor cell output in humans and three nonhuman primates using directed differentiation of pluripotent stem cells (PSCs) in adherent two-dimensional (2D) and organoid three-dimensional (3D) culture systems. Clonal lineage analysis showed that primate cortical progenitors proliferate for a protracted period of time, during which they generate early-born neurons, in contrast to rodents, where this expansion phase largely ceases before neurogenesis begins. The extent of this additional cortical progenitor expansion differs among primates, leading to differences in the number of neurons generated by each progenitor cell. We found that this mechanism for controlling cortical size is regulated cell autonomously in culture, suggesting that primate cerebral cortex size is regulated at least in part at the level of individual cortical progenitor cell clonal output.
Copyright © 2016 The Authors. Published by Elsevier Inc. All rights reserved.
Figures

Comment in
-
Stem Cells Expand Insights into Human Brain Evolution.Cell Stem Cell. 2016 Apr 7;18(4):425-6. doi: 10.1016/j.stem.2016.03.017. Cell Stem Cell. 2016. PMID: 27058930
-
From stem cells to comparative corticogenesis: a bridge too far?Stem Cell Investig. 2016 Aug 16;3:39. doi: 10.21037/sci.2016.08.02. eCollection 2016. Stem Cell Investig. 2016. PMID: 27668246 Free PMC article.
References
-
- Azevedo F.A.C., Carvalho L.R.B., Grinberg L.T., Farfel J.M., Ferretti R.E.L., Leite R.E.P., Jacob Filho W., Lent R., Herculano-Houzel S. Equal numbers of neuronal and nonneuronal cells make the human brain an isometrically scaled-up primate brain. J. Comp. Neurol. 2009;513:532–541. - PubMed
-
- Dehay C., Kennedy H., Kosik K.S. The outer subventricular zone and primate-specific cortical complexification. Neuron. 2015;85:683–694. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
