

TAM receptors regulate multiple features of microglial physiology
- PMID: 27049947
- PMCID: PMC5358512
- DOI: 10.1038/nature17630
TAM receptors regulate multiple features of microglial physiology
Abstract
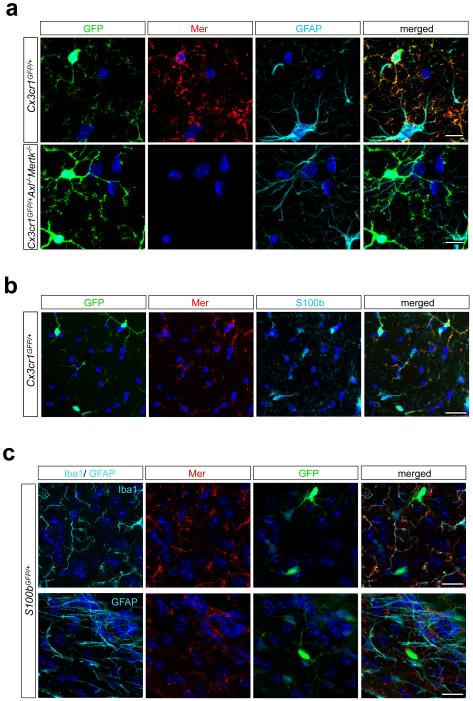
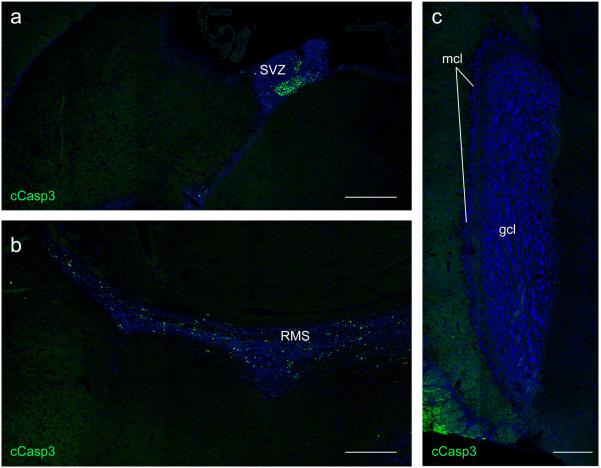
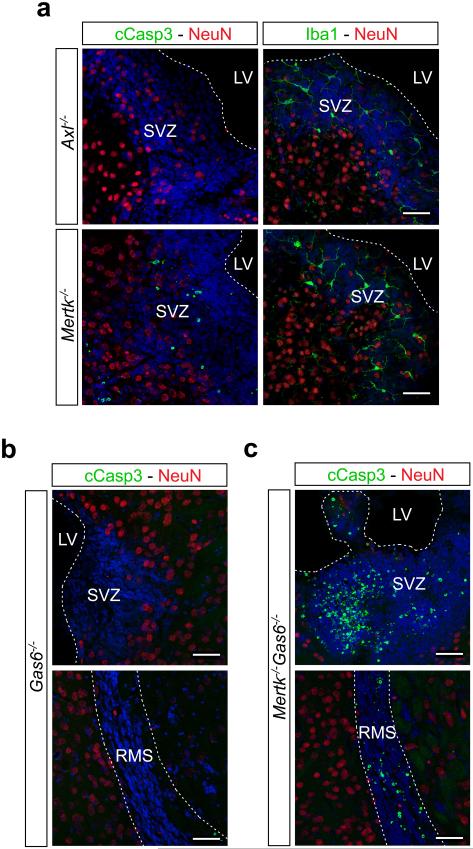
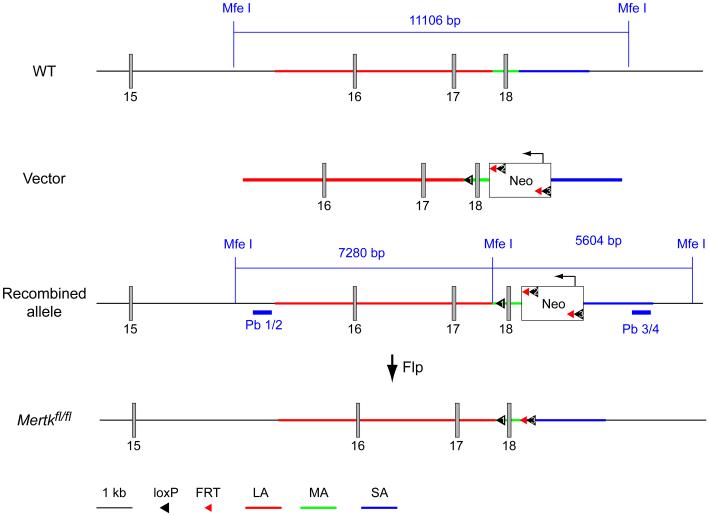
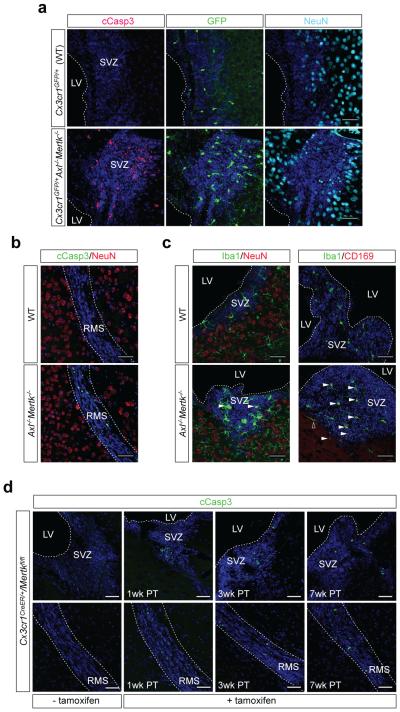
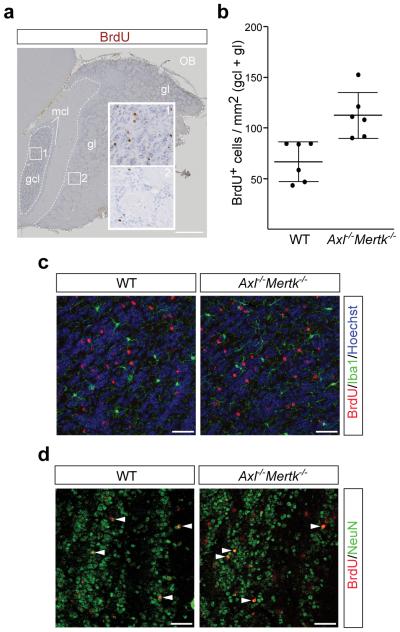
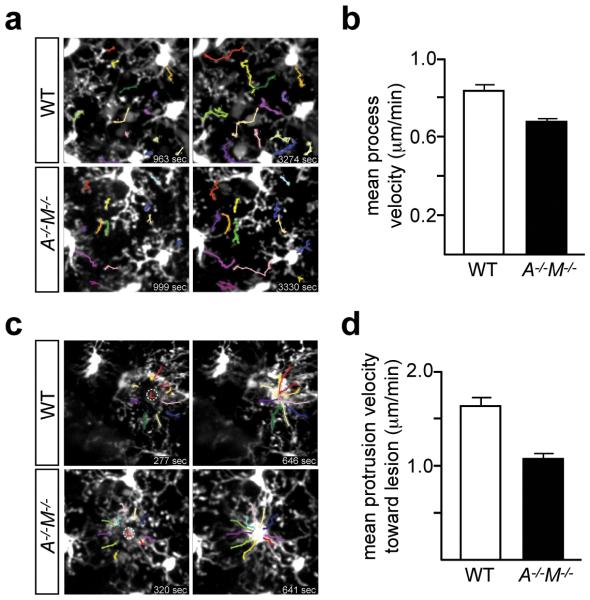
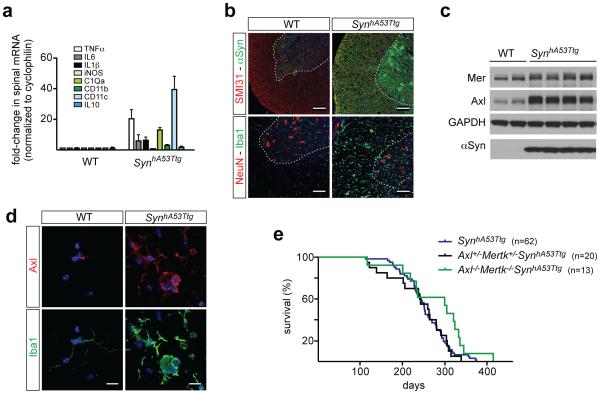
Microglia are damage sensors for the central nervous system (CNS), and the phagocytes responsible for routine non-inflammatory clearance of dead brain cells. Here we show that the TAM receptor tyrosine kinases Mer and Axl regulate these microglial functions. We find that adult mice deficient in microglial Mer and Axl exhibit a marked accumulation of apoptotic cells specifically in neurogenic regions of the CNS, and that microglial phagocytosis of the apoptotic cells generated during adult neurogenesis is normally driven by both TAM receptor ligands Gas6 and protein S. Using live two-photon imaging, we demonstrate that the microglial response to brain damage is also TAM-regulated, as TAM-deficient microglia display reduced process motility and delayed convergence to sites of injury. Finally, we show that microglial expression of Axl is prominently upregulated in the inflammatory environment that develops in a mouse model of Parkinson's disease. Together, these results establish TAM receptors as both controllers of microglial physiology and potential targets for therapeutic intervention in CNS disease.
Figures
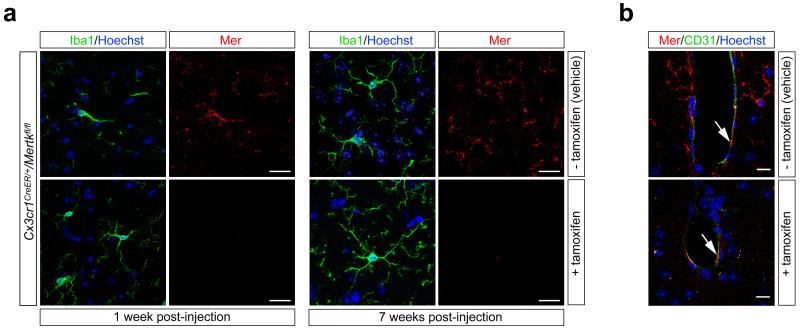
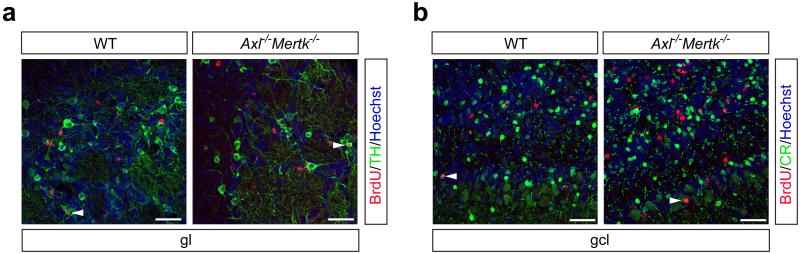
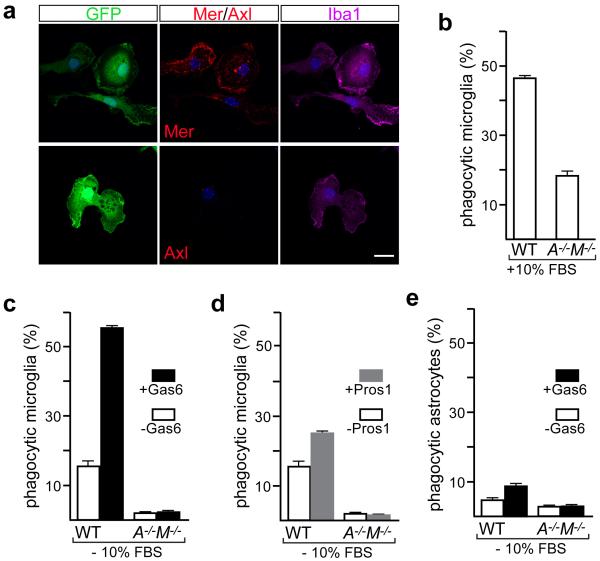
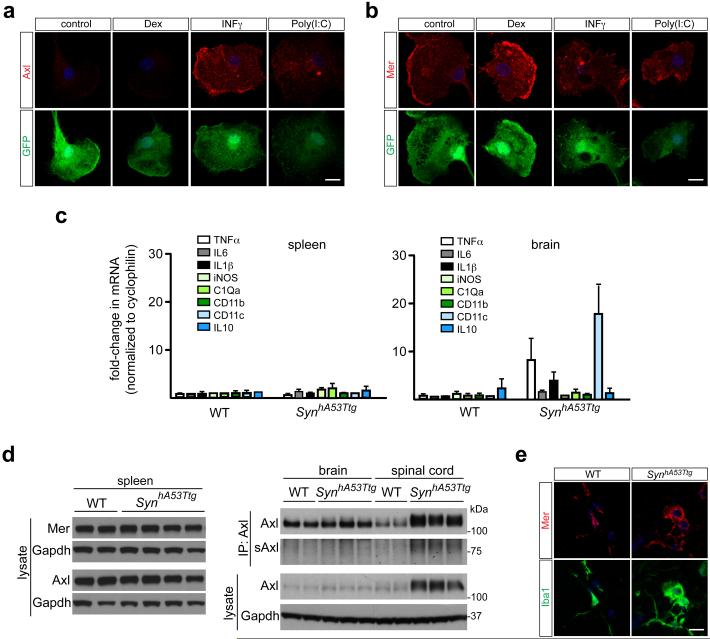
Comment in
-
Neuroinflammation: Surprises from the sanitary engineers.Nature. 2016 Apr 14;532(7598):185-6. doi: 10.1038/nature17881. Epub 2016 Apr 6. Nature. 2016. PMID: 27049948 No abstract available.
References
-
- Ransohoff RM, Cardona AE. The myeloid cells of the central nervous system parenchyma. Nature. 2010;468:253–262. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
