

Differential Mitochondrial Requirements for Radially and Non-radially Migrating Cortical Neurons: Implications for Mitochondrial Disorders
- PMID: 27050514
- PMCID: PMC5412722
- DOI: 10.1016/j.celrep.2016.03.024
Differential Mitochondrial Requirements for Radially and Non-radially Migrating Cortical Neurons: Implications for Mitochondrial Disorders
Abstract
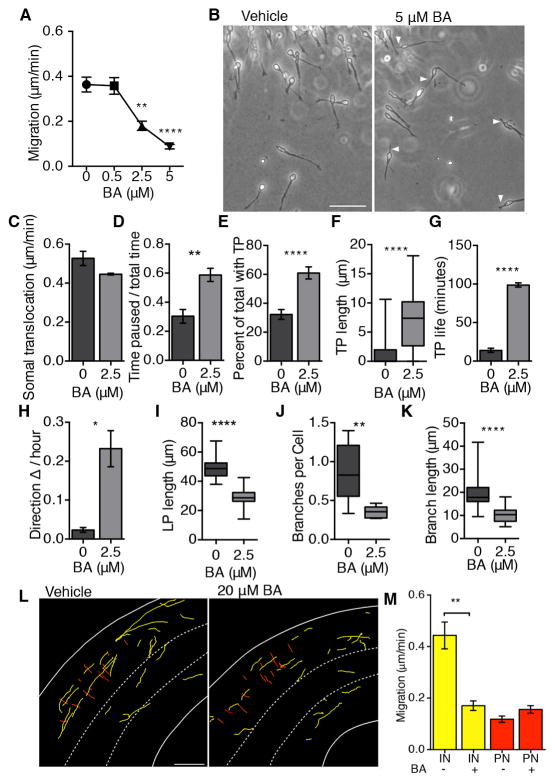
Mitochondrial dysfunction has been increasingly linked to neurodevelopmental disorders such as intellectual disability, childhood epilepsy, and autism spectrum disorder, conditions also associated with cortical GABAergic interneuron dysfunction. Although interneurons have some of the highest metabolic demands in the postnatal brain, the importance of mitochondria during interneuron development is unknown. We find that interneuron migration from the basal forebrain to the neocortex is highly sensitive to perturbations in oxidative phosphorylation. Both pharmacologic and genetic inhibition of adenine nucleotide transferase 1 (Ant1) disrupts the non-radial migration of interneurons, but not the radial migration of cortical projection neurons. The selective dependence of cortical interneuron migration on oxidative phosphorylation may be a mechanistic pathway upon which multiple developmental and metabolic pathologies converge.
Copyright © 2016 The Authors. Published by Elsevier Inc. All rights reserved.
Figures




Similar articles
-
Interneuron development and dysfunction.FEBS J. 2022 Apr;289(8):2318-2336. doi: 10.1111/febs.15872. Epub 2021 Apr 23. FEBS J. 2022. PMID: 33844440 Review.
-
Interneuron odyssey: molecular mechanisms of tangential migration.Front Neural Circuits. 2023 Sep 14;17:1256455. doi: 10.3389/fncir.2023.1256455. eCollection 2023. Front Neural Circuits. 2023. PMID: 37779671 Free PMC article. Review.
-
Cxcr4 regulation of interneuron migration is disrupted in 22q11.2 deletion syndrome.Proc Natl Acad Sci U S A. 2012 Nov 6;109(45):18601-6. doi: 10.1073/pnas.1211507109. Epub 2012 Oct 22. Proc Natl Acad Sci U S A. 2012. PMID: 23091025 Free PMC article.
-
Control of tangential/non-radial migration of neurons in the developing cerebral cortex.Neurochem Int. 2007 Jul-Sep;51(2-4):121-31. doi: 10.1016/j.neuint.2007.05.006. Epub 2007 May 21. Neurochem Int. 2007. PMID: 17588709 Review.
-
Pax6-/- mice have a cell nonautonomous defect in nonradial interneuron migration.Cereb Cortex. 2008 Apr;18(4):752-62. doi: 10.1093/cercor/bhm114. Epub 2007 Jul 18. Cereb Cortex. 2008. PMID: 17634386
Cited by
-
Cellular stress mechanisms of prenatal maternal stress: Heat shock factors and oxidative stress.Neurosci Lett. 2019 Sep 14;709:134368. doi: 10.1016/j.neulet.2019.134368. Epub 2019 Jul 9. Neurosci Lett. 2019. PMID: 31299286 Free PMC article. Review.
-
Miro1-dependent mitochondrial dynamics in parvalbumin interneurons.Elife. 2021 Jun 30;10:e65215. doi: 10.7554/eLife.65215. Elife. 2021. PMID: 34190042 Free PMC article.
-
Maternal Inflammation During Pregnancy and Offspring Brain Development: The Role of Mitochondria.Biol Psychiatry Cogn Neurosci Neuroimaging. 2022 May;7(5):498-509. doi: 10.1016/j.bpsc.2021.11.003. Epub 2021 Nov 17. Biol Psychiatry Cogn Neurosci Neuroimaging. 2022. PMID: 34800727 Free PMC article. Review.
-
Integrated RNA Sequencing Reveals Epigenetic Impacts of Diesel Particulate Matter Exposure in Human Cerebral Organoids.Dev Neurosci. 2020;42(5-6):195-207. doi: 10.1159/000513536. Epub 2021 Mar 3. Dev Neurosci. 2020. PMID: 33657557 Free PMC article.
-
FARS2 mutations presenting with pure spastic paraplegia and lesions of the dentate nuclei.Ann Clin Transl Neurol. 2018 Aug 14;5(9):1128-1133. doi: 10.1002/acn3.598. eCollection 2018 Sep. Ann Clin Transl Neurol. 2018. PMID: 30250868 Free PMC article.
References
-
- Adeva-Andany M, Lopez-Ojen M, Funcasta-Calderon R, Ameneiros-Rodriguez E, Donapetry-Garcia C, Vila-Altesor M, Rodriguez-Seijas J. Comprehensive review on lactate metabolism in human health. Mitochondrion. 2014;17C:76–100. - PubMed
-
- Bellion A, Baudoin JP, Alvarez C, Bornens M, Metin C. Nucleokinesis in tangentially migrating neurons comprises two alternating phases: forward migration of the Golgi/centrosome associated with centrosome splitting and myosin contraction at the rear. The Journal of neuroscience: the official journal of the Society for Neuroscience. 2005;25:5691–5699. - PMC - PubMed
-
- Benson DL, Watkins FH, Steward O, Banker G. Characterization of GABAergic neurons in hippocampal cell cultures. Journal of neurocytology. 1994;23:279–295. - PubMed
-
- Bertholet AM, Millet AM, Guillermin O, Daloyau M, Davezac N, Miquel MC, Belenguer P. OPA1 loss of function affects in vitro neuronal maturation. Brain: a journal of neurology. 2013;136:1518–1533. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
