

PRRT2 Is a Key Component of the Ca(2+)-Dependent Neurotransmitter Release Machinery
- PMID: 27052163
- PMCID: PMC4826441
- DOI: 10.1016/j.celrep.2016.03.005
PRRT2 Is a Key Component of the Ca(2+)-Dependent Neurotransmitter Release Machinery
Abstract
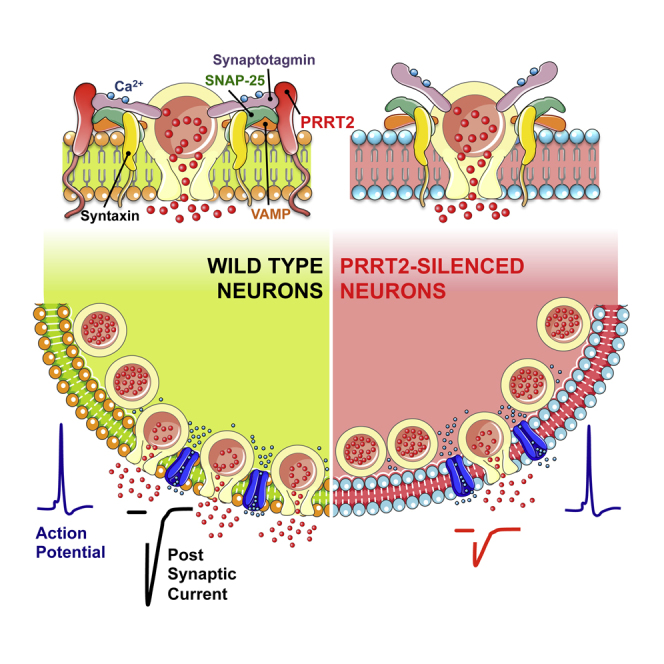
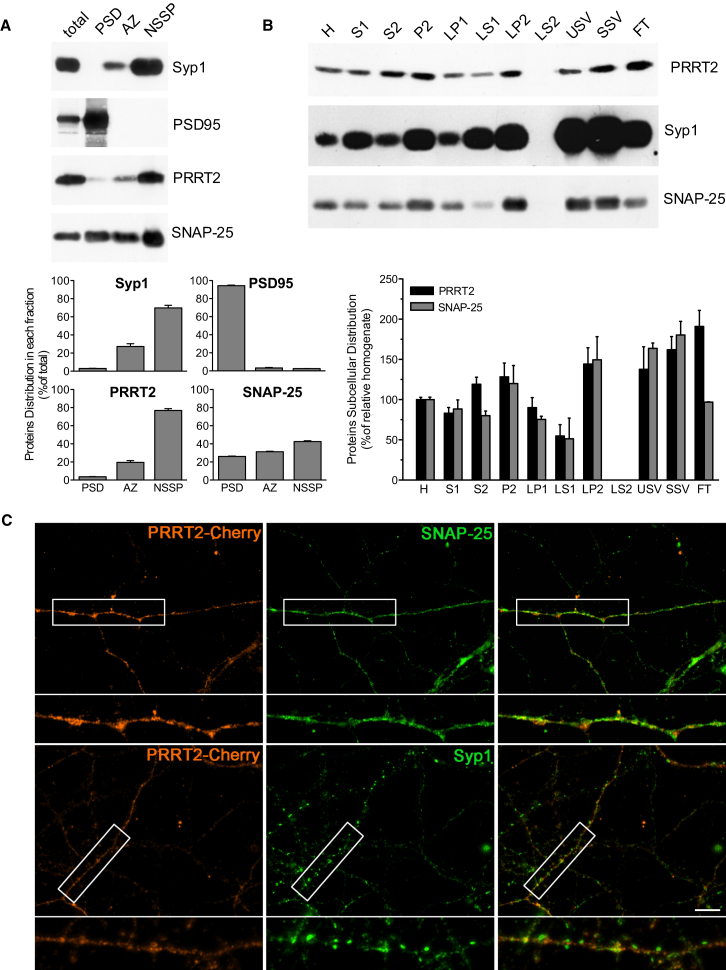
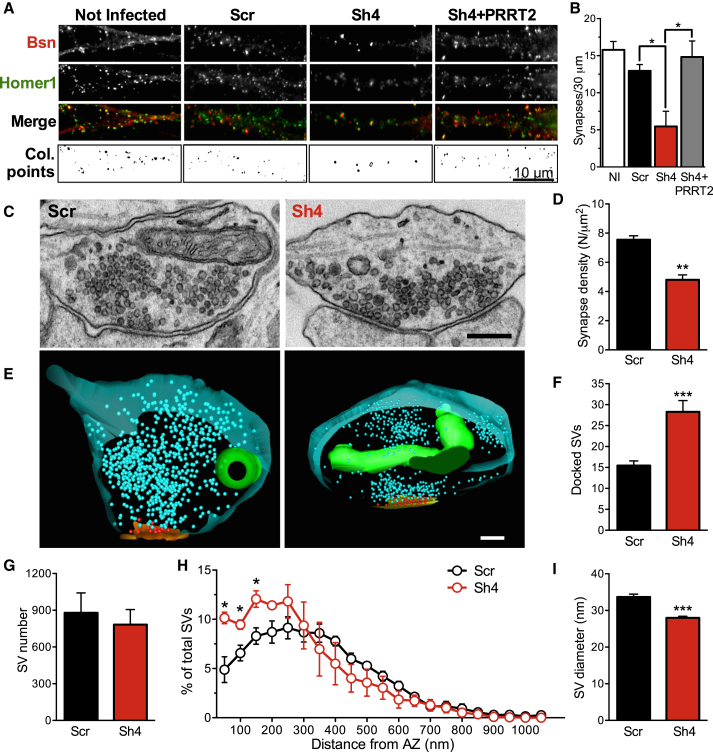
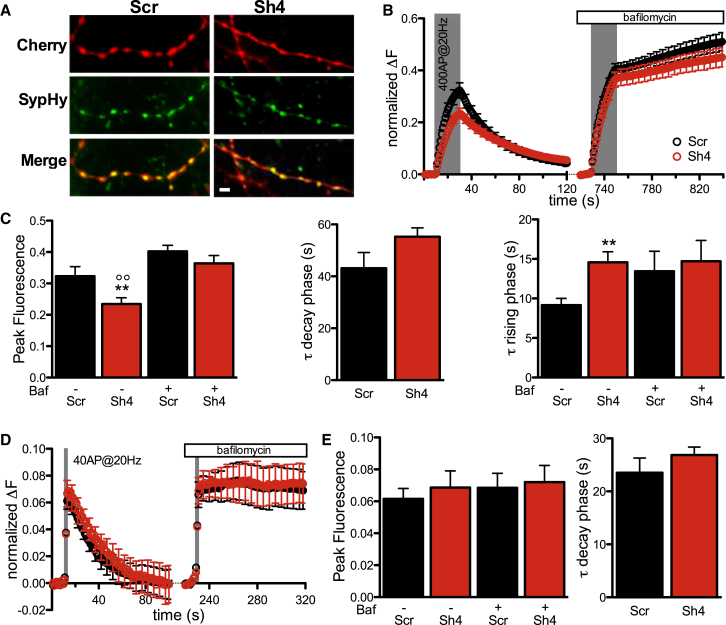
Heterozygous mutations in proline-rich transmembrane protein 2 (PRRT2) underlie a group of paroxysmal disorders, including epilepsy, kinesigenic dyskinesia, and migraine. Most of the mutations lead to impaired PRRT2 expression, suggesting that loss of PRRT2 function may contribute to pathogenesis. We show that PRRT2 is enriched in presynaptic terminals and that its silencing decreases the number of synapses and increases the number of docked synaptic vesicles at rest. PRRT2-silenced neurons exhibit a severe impairment of synchronous release, attributable to a sharp decrease in release probability and Ca(2+) sensitivity and associated with a marked increase of the asynchronous/synchronous release ratio. PRRT2 interacts with the synaptic proteins SNAP-25 and synaptotagmin 1/2. The results indicate that PRRT2 is intimately connected with the Ca(2+)-sensing machinery and that it plays an important role in the final steps of neurotransmitter release.
Keywords: PRRT2; knockdown; release probability; synaptic transmission; synaptotagmin; synchronous and asynchronous release.
Copyright © 2016 The Authors. Published by Elsevier Inc. All rights reserved.
Figures
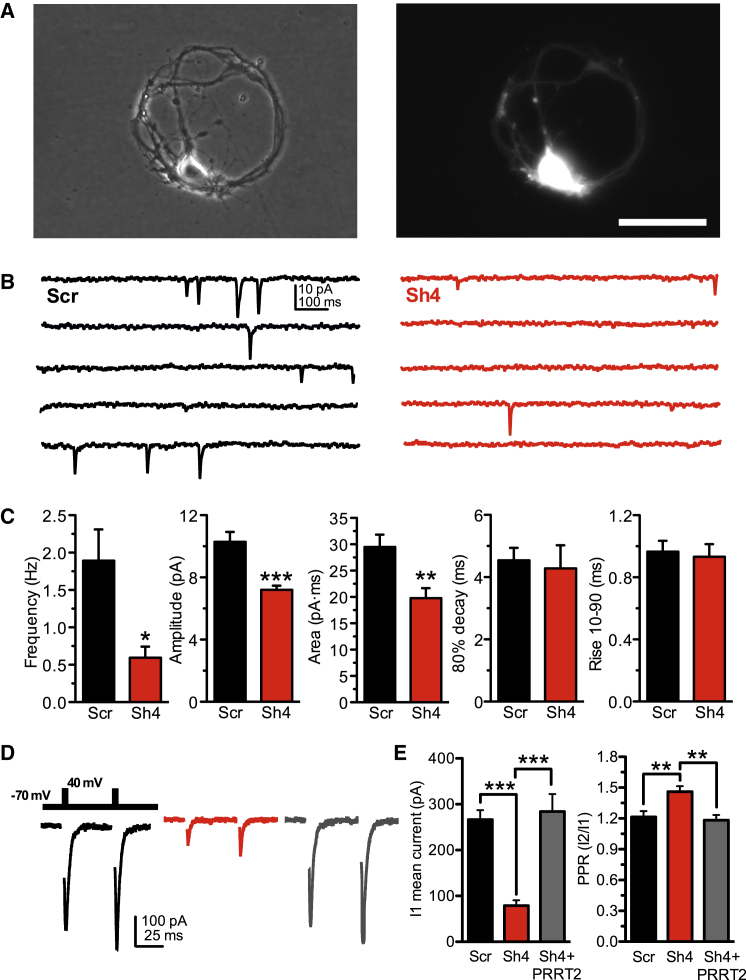
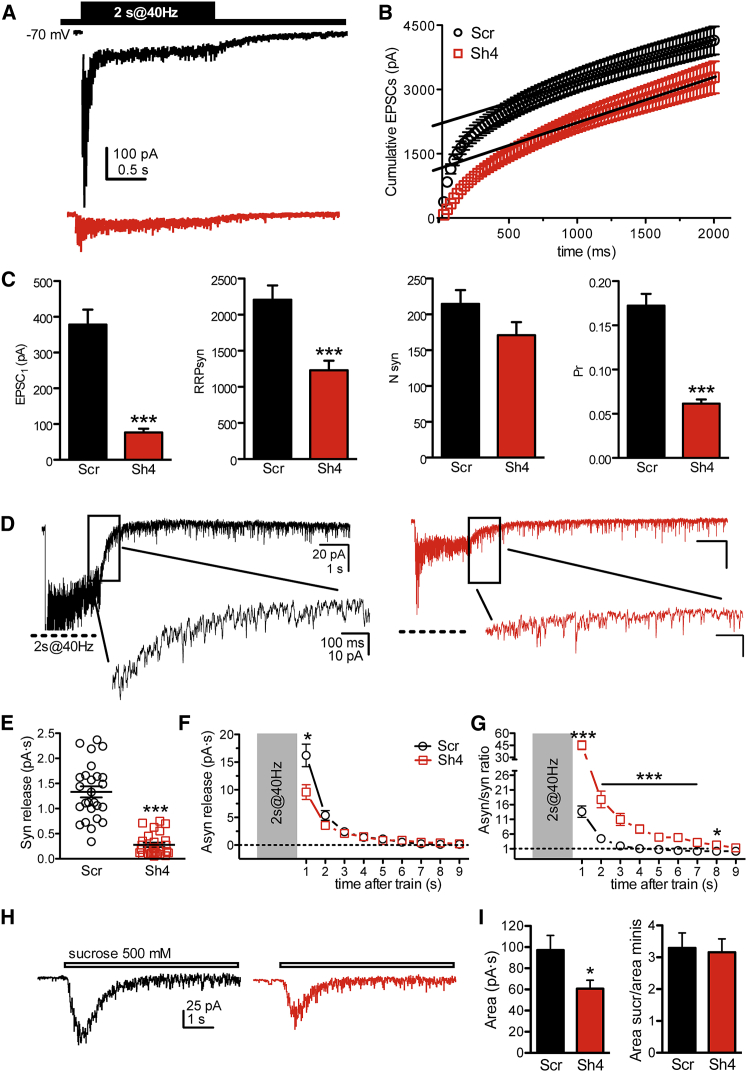
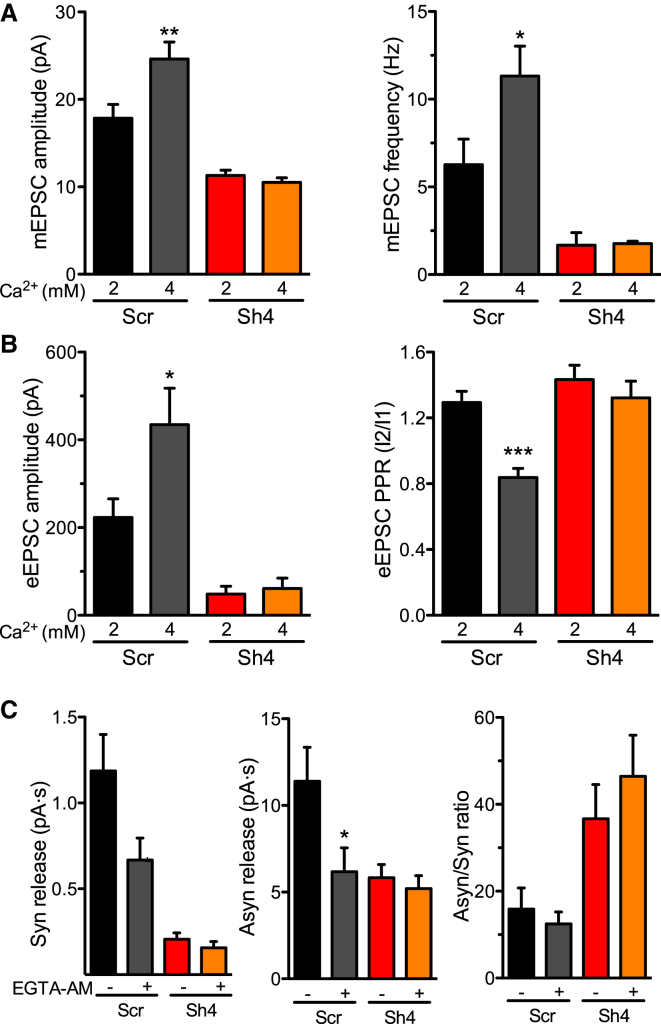
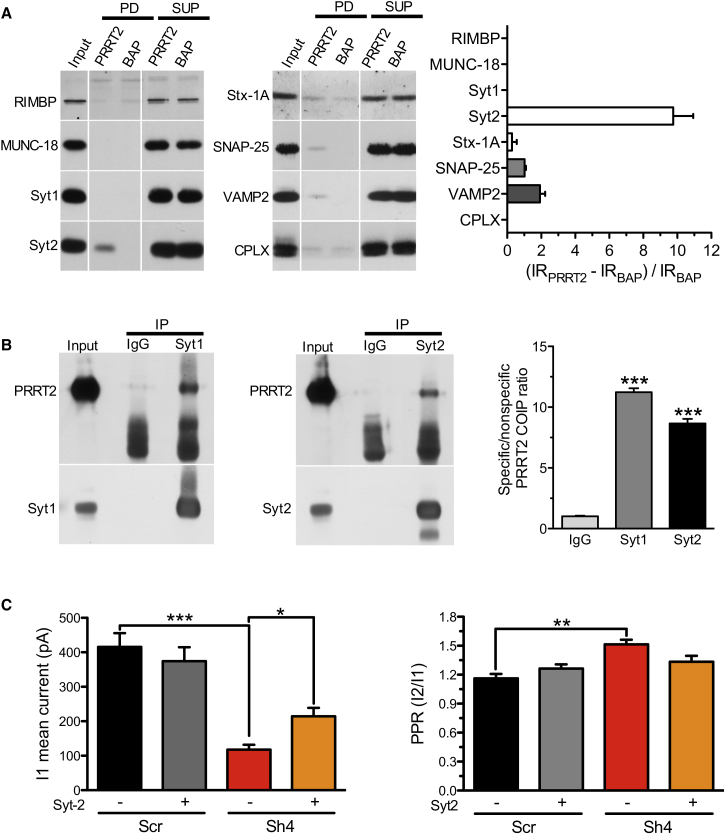
Comment in
-
The Role of PRRT2 in Synaptic Transmission May Not Be So Benign.Epilepsy Curr. 2017 May-Jun;17(3):165-166. doi: 10.5698/1535-7511.17.3.165. Epilepsy Curr. 2017. PMID: 28684951 Free PMC article. No abstract available.
References
-
- Chen W.J., Lin Y., Xiong Z.Q., Wei W., Ni W., Tan G.H., Guo S.L., He J., Chen Y.F., Zhang Q.J. Exome sequencing identifies truncating mutations in PRRT2 that cause paroxysmal kinesigenic dyskinesia. Nat. Genet. 2011;43:1252–1255. - PubMed
-
- Chiappalone M., Casagrande S., Tedesco M., Valtorta F., Baldelli P., Martinoia S., Benfenati F. Opposite changes in glutamatergic and GABAergic transmission underlie the diffuse hyperexcitability of synapsin I-deficient cortical networks. Cereb. Cortex. 2009;19:1422–1439. - PubMed
-
- Ebrahimi-Fakhari D., Saffari A., Westenberger A., Klein C. The evolving spectrum of PRRT2-associated paroxysmal diseases. Brain. 2015;138:3476–3495. - PubMed
-
- Fox M.A., Sanes J.R. Synaptotagmin I and II are present in distinct subsets of central synapses. J. Comp. Neurol. 2007;503:280–296. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
