

Multiple sites of adaptation lead to contrast encoding in the Drosophila olfactory system
- PMID: 27053295
- PMCID: PMC4831330
- DOI: 10.14814/phy2.12762
Multiple sites of adaptation lead to contrast encoding in the Drosophila olfactory system
Abstract
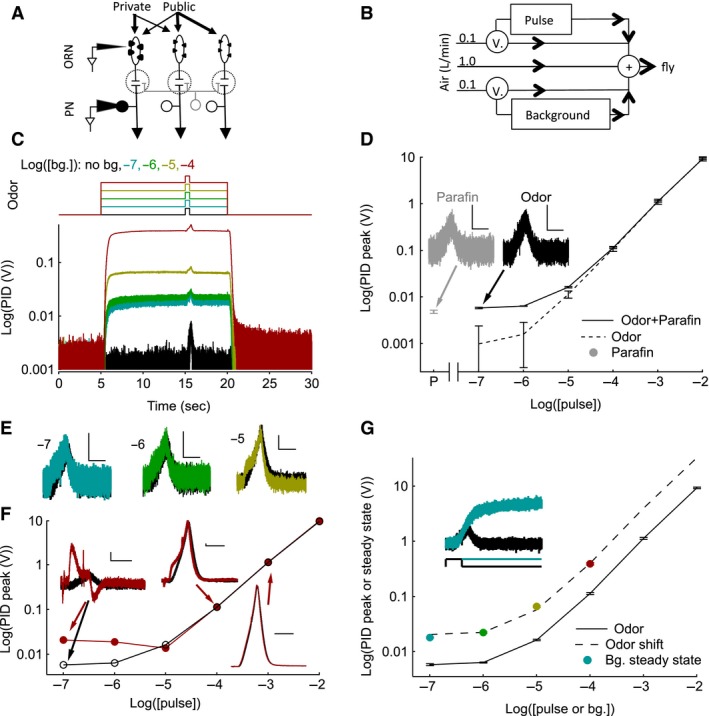
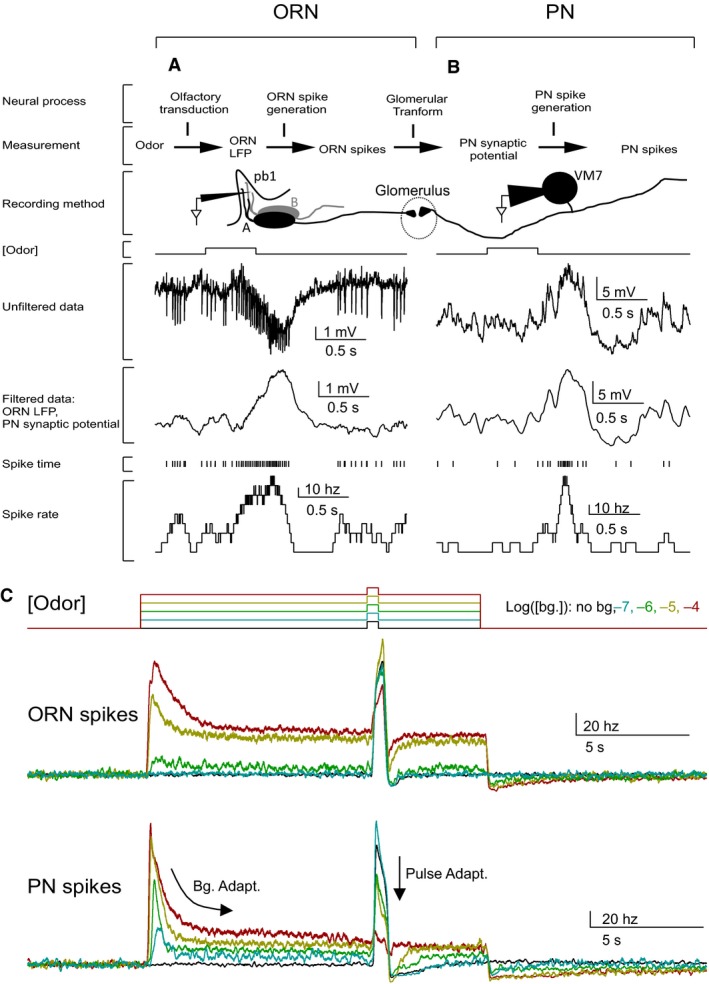
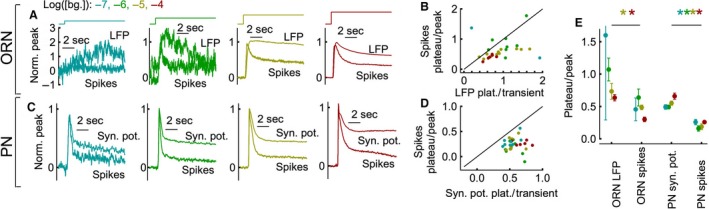
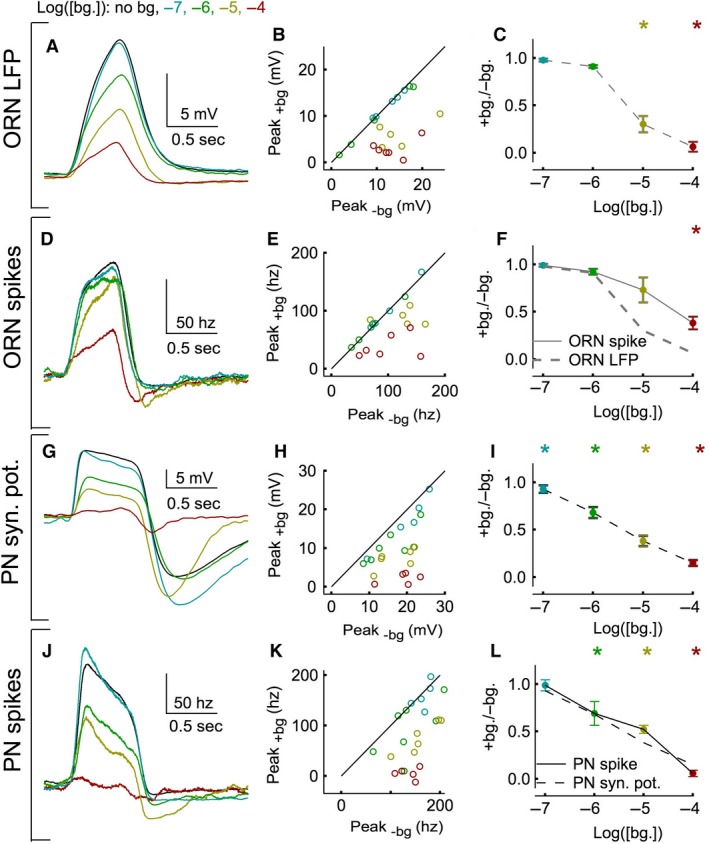
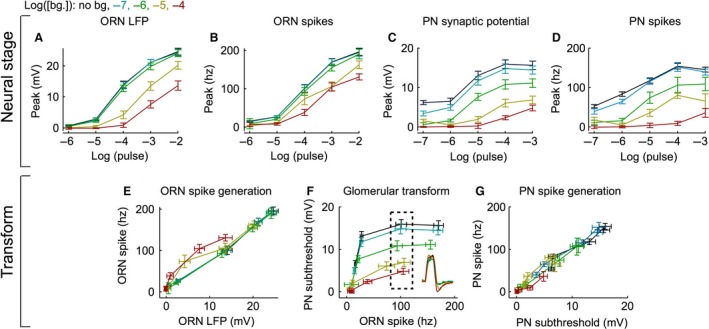
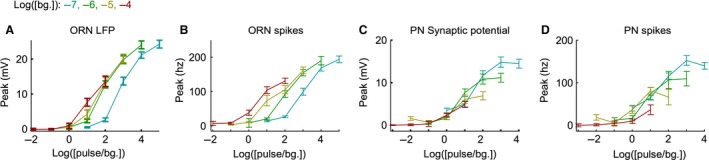
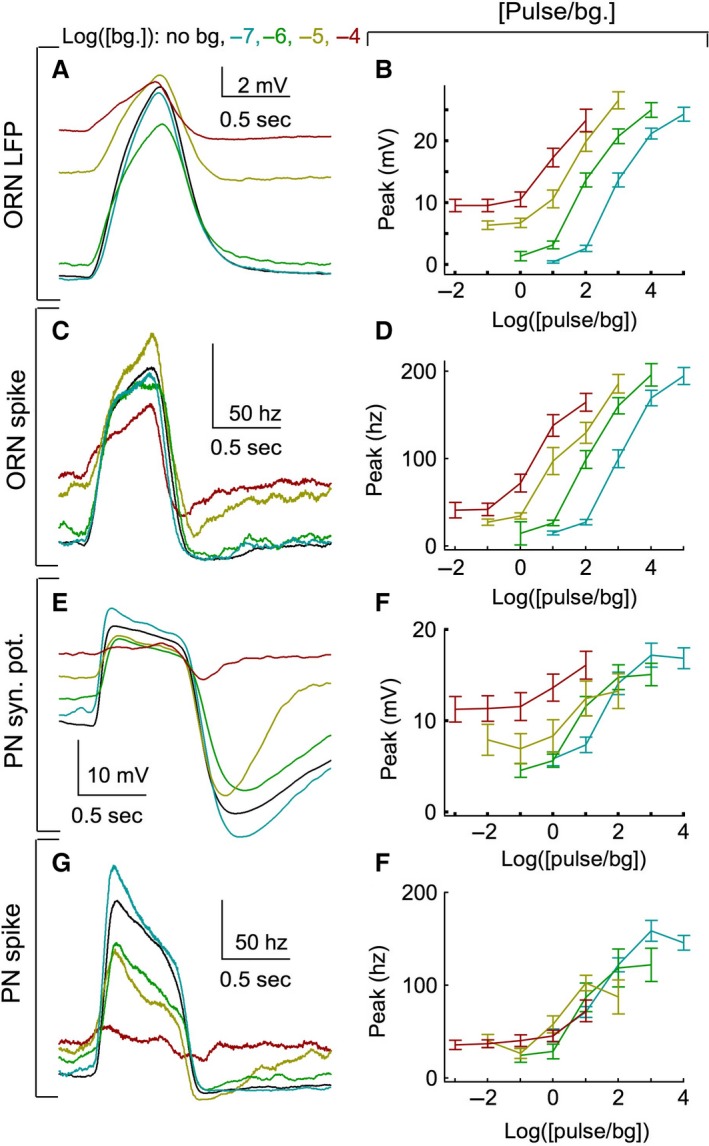
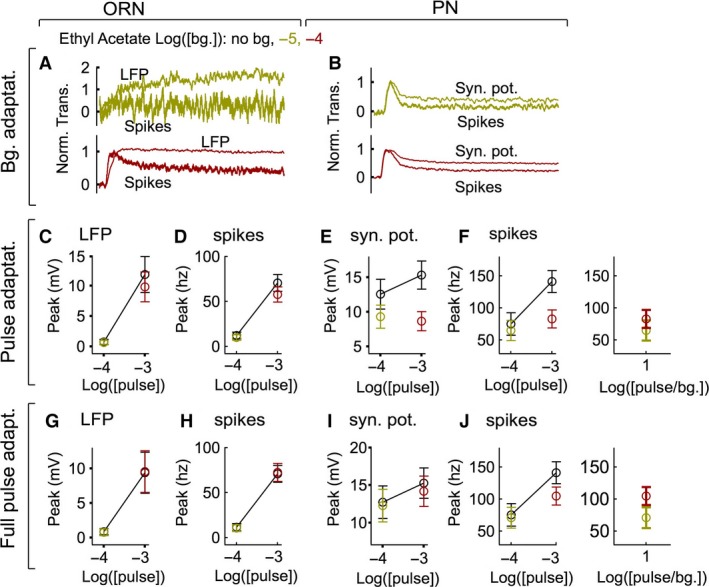
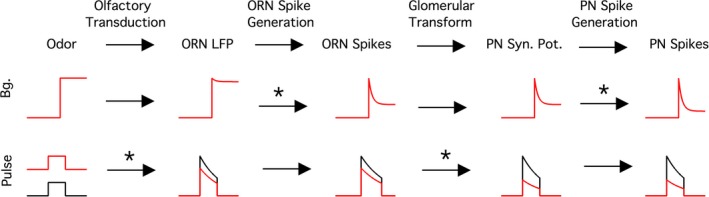
Animals often encounter large increases in odor intensity that can persist for many seconds. These increases in the background odor are often accompanied by increases in the variance of the odor stimulus. Previous studies have shown that a persistent odor stimulus (odor background) results in a decrease in the response to brief odor pulses in the olfactory receptor neurons (ORNs). However, the contribution of adapting mechanisms beyond theORNs is not clear. Thus, it is unclear how adaptive mechanisms are distributed within the olfactory circuit and what impact downstream adaptation may have on the encoding of odor stimuli. In this study, adaptation to the same odor stimulus is examined at multiple levels in the well studied and accessibleDrosophilaolfactory system. The responses of theORNs are compared to the responses of the second order, projection neurons (PNs), directly connected to them. Adaptation inPNspike rate was found to be much greater than adaptation in theORNspike rate. This greater adaptation allowsPNs to encode odor contrast (ratio of pulse intensity to background intensity) with little ambiguity. Moreover, distinct neural mechanisms contribute to different aspects of adaptation; adaptation to the background odor is dominated by adaptation in spike generation in bothORNs andPNs, while adaptation to the odor pulse is dominated by changes within olfactory transduction and the glomerulus. These observations suggest that the olfactory system adapts at multiple sites to better match its response gain to stimulus statistics.
Keywords: Adaptation; Drosophila; Olfaction; Weber's law.
© 2016 The Authors. Physiological Reports published by Wiley Periodicals, Inc. on behalf of the American Physiological Society and The Physiological Society.
Figures
References
-
- Barlow, H. B. 1961. Possible principles underlying the transformations of sensory messages Pp. 217–234 in Rosenblith W., ed. Sensory communication. Ch13. M.I.T. Press, Cambridge, MA:
-
- Brown, S. L. , Joseph J., and Stopfer M.. 2005. Encoding a temporally structured stimulus with a temporally structured neural representation. Nat. Neurosci. 8:1568–1576. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
