

Antibody Responses with Fc-Mediated Functions after Vaccination of HIV-Infected Subjects with Trivalent Influenza Vaccine
- PMID: 27053553
- PMCID: PMC4886795
- DOI: 10.1128/JVI.00285-16
Antibody Responses with Fc-Mediated Functions after Vaccination of HIV-Infected Subjects with Trivalent Influenza Vaccine
Abstract
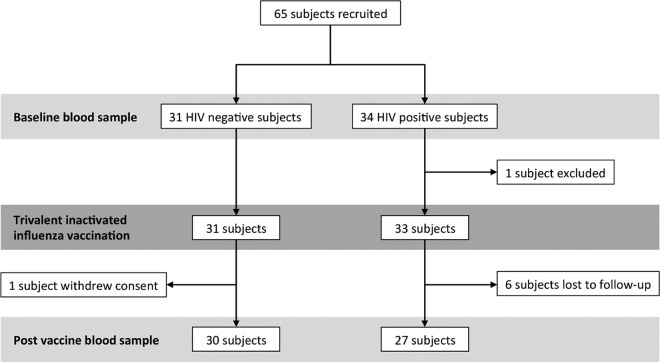
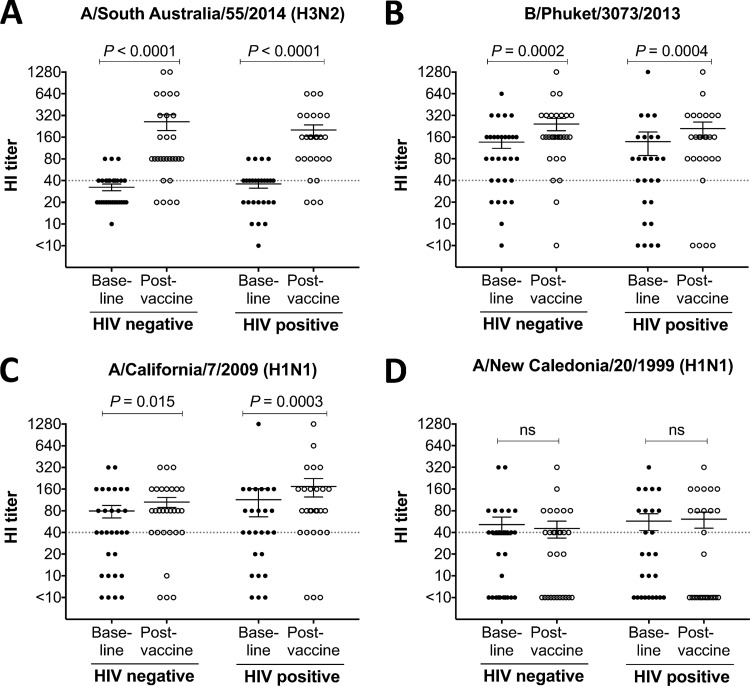
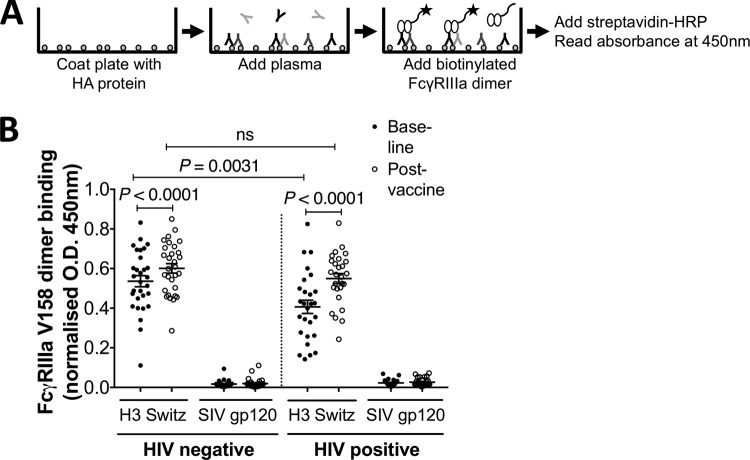
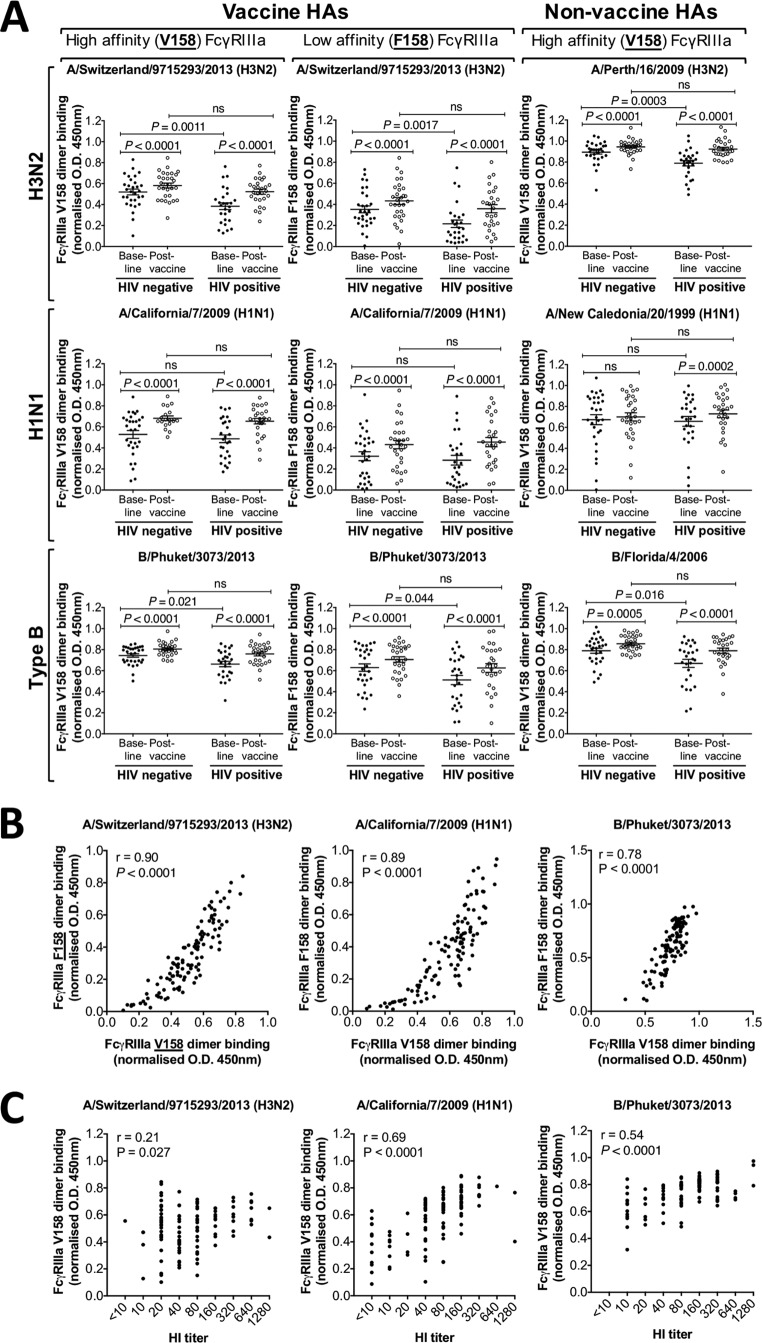
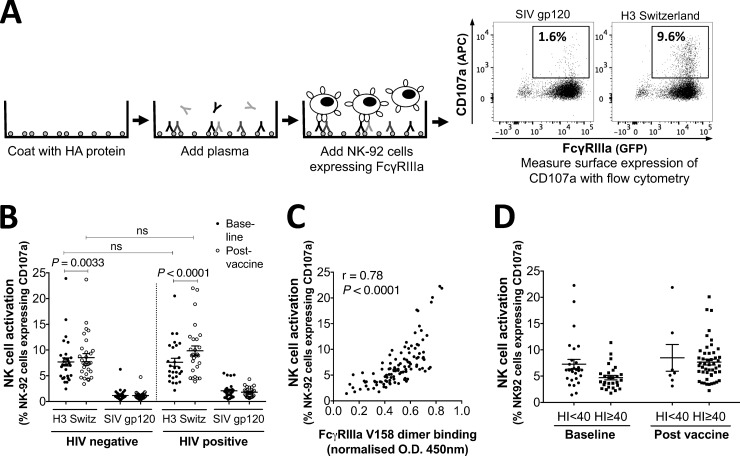
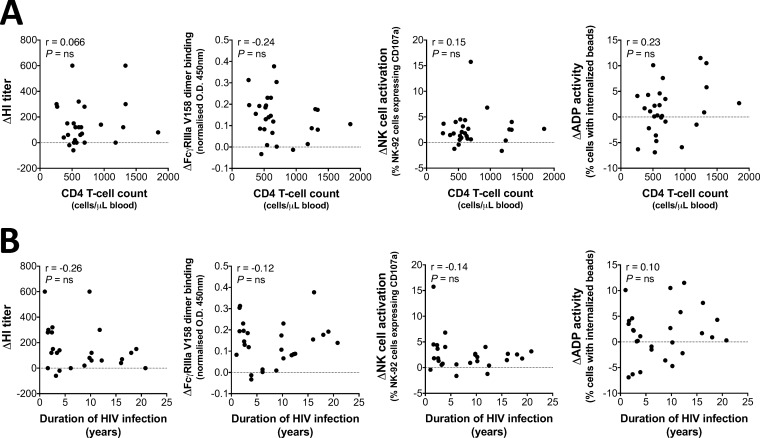
This study seeks to assess the ability of seasonal trivalent inactivated influenza vaccine (TIV) to induce nonneutralizing antibodies (Abs) with Fc-mediated functions in HIV-uninfected and HIV-infected subjects. Functional influenza-specific Ab responses were studied in 30 HIV-negative and 27 HIV-positive subjects immunized against seasonal influenza. All 57 subjects received the 2015 TIV. Fc-mediated antihemagglutinin (anti-HA) Ab activity was measured in plasma before and 4 weeks after vaccination using Fc-receptor-binding assays, NK cell activation assays, and phagocytosis assays. At baseline, the HIV-positive group had detectable but reduced functional Ab responses to both vaccine and nonvaccine influenza antigens. TIV enhanced Fc-mediated Ab responses in both HIV-positive and HIV-negative groups. A larger rise was generally observed in the HIV-positive group, such that there was no difference in functional Ab responses between the two groups after vaccination. The 2015 TIV enhanced functional influenza-specific Ab responses in both HIV-negative and HIV-positive subjects to a range of influenza HA proteins. The increase in functional Ab responses in the HIV-positive group supports recommendations to immunize this at-risk group.
Importance: Infection with HIV is associated with increasing disease severity following influenza infections, and annual influenza vaccinations are recommended for this target group. However, HIV-infected individuals respond relatively poorly to vaccination compared to healthy individuals, particularly if immunodeficient. There is therefore a need to increase our understanding of immunity to influenza in the context of underlying HIV infection. While antibodies can mediate direct virus neutralization, interactions with cellular Fc receptors may be important for anti-influenza immunity in vivo by facilitating antibody-dependent cellular cytotoxicity (ADCC) and/or antibody-dependent phagocytosis (ADP). The ability of seasonal influenza vaccines to induce antibody responses with potent Fc-mediated antiviral activity is currently unclear. Probing the ADCC and ADP responses to influenza vaccination has provided important new information in the quest to improve immunity to influenza.
Copyright © 2016, American Society for Microbiology. All Rights Reserved.
Figures

Similar articles
-
Hemagglutinin Stalk-Specific Fc-Mediated Functions Are Associated With Protection Against Influenza Illness After Seasonal Influenza Vaccination.J Infect Dis. 2024 Dec 16;230(6):1329-1336. doi: 10.1093/infdis/jiae241. J Infect Dis. 2024. PMID: 38743692 Free PMC article. Clinical Trial.
-
Antibody-dependent phagocytosis (ADP) responses following trivalent inactivated influenza vaccination of younger and older adults.Vaccine. 2017 Nov 7;35(47):6451-6458. doi: 10.1016/j.vaccine.2017.09.062. Epub 2017 Oct 10. Vaccine. 2017. PMID: 29029940
-
Standard trivalent influenza virus protein vaccination does not prime antibody-dependent cellular cytotoxicity in macaques.J Virol. 2013 Dec;87(24):13706-18. doi: 10.1128/JVI.01666-13. Epub 2013 Oct 9. J Virol. 2013. PMID: 24109221 Free PMC article.
-
Extra-Neutralizing FcR-Mediated Antibody Functions for a Universal Influenza Vaccine.Front Immunol. 2019 Mar 18;10:440. doi: 10.3389/fimmu.2019.00440. eCollection 2019. Front Immunol. 2019. PMID: 30949165 Free PMC article. Review.
-
Fc or not Fc; that is the question: Antibody Fc-receptor interactions are key to universal influenza vaccine design.Hum Vaccin Immunother. 2017 Jun 3;13(6):1-9. doi: 10.1080/21645515.2017.1290018. Epub 2017 Mar 23. Hum Vaccin Immunother. 2017. PMID: 28332900 Free PMC article. Review.
Cited by
-
A Method to Assess Fc-mediated Effector Functions Induced by Influenza Hemagglutinin Specific Antibodies.J Vis Exp. 2018 Feb 23;(132):56256. doi: 10.3791/56256. J Vis Exp. 2018. PMID: 29553549 Free PMC article.
-
Antibody-dependent NK cell degranulation as a marker for assessing antibody-dependent cytotoxicity against pandemic 2009 influenza A(H1N1) infection in human plasma and influenza-vaccinated transchromosomic bovine intravenous immunoglobulin therapy.J Virol Methods. 2017 Oct;248:7-18. doi: 10.1016/j.jviromet.2017.06.007. Epub 2017 Jun 15. J Virol Methods. 2017. PMID: 28624584 Free PMC article.
-
Brief Report: Decreased JC Virus-Specific Antibody-Dependent Cellular Cytotoxicity in HIV-Seropositive PML Survivors.J Acquir Immune Defic Syndr. 2019 Oct 1;82(2):220-224. doi: 10.1097/QAI.0000000000002105. J Acquir Immune Defic Syndr. 2019. PMID: 31513076 Free PMC article.
-
The Inhibition of H1N1 Influenza Virus-Induced Apoptosis by Surface Decoration of Selenium Nanoparticles with β-Thujaplicin through Reactive Oxygen Species-Mediated AKT and p53 Signaling Pathways.ACS Omega. 2020 Nov 16;5(47):30633-30642. doi: 10.1021/acsomega.0c04624. eCollection 2020 Dec 1. ACS Omega. 2020. PMID: 33283112 Free PMC article.
-
Antibody mediated activation of natural killer cells in malaria exposed pregnant women.Sci Rep. 2021 Feb 18;11(1):4130. doi: 10.1038/s41598-021-83093-4. Sci Rep. 2021. PMID: 33602987 Free PMC article.
References
-
- Cohen C, Moyes J, Tempia S, Groom M, Walaza S, Pretorius M, Dawood H, Chhagan M, Haffejee S, Variava E, Kahn K, Tshangela A, von Gottberg A, Wolter N, Cohen AL, Kgokong B, Venter M, Madhi SA. 2013. Severe influenza-associated respiratory infection in high HIV prevalence setting, South Africa, 2009-2011. Emerg Infect Dis 19:1766–1774. doi:10.3201/eid1911.130546. - DOI - PMC - PubMed
-
- Tasker SA, Treanor JJ, Paxton WB, Wallace MR. 1999. Efficacy of influenza vaccination in HIV-infected persons. A randomized, double-blind, placebo-controlled trial. Ann Intern Med 131:430–433. - PubMed
-
- Wang TT, Tan GS, Hai R, Pica N, Ngai L, Ekiert DC, Wilson IA, Garcia-Sastre A, Moran TM, Palese P. 2010. Vaccination with a synthetic peptide from the influenza virus hemagglutinin provides protection against distinct viral subtypes. Proc Natl Acad Sci U S A 107:18979–18984. doi:10.1073/pnas.1013387107. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
