

How Soluble GARP Enhances TGFβ Activation
- PMID: 27054568
- PMCID: PMC4824412
- DOI: 10.1371/journal.pone.0153290
How Soluble GARP Enhances TGFβ Activation
Abstract
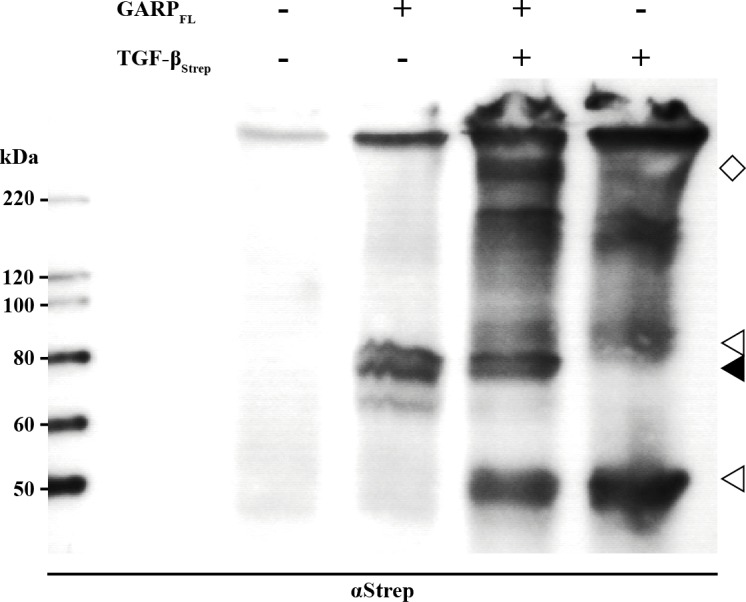
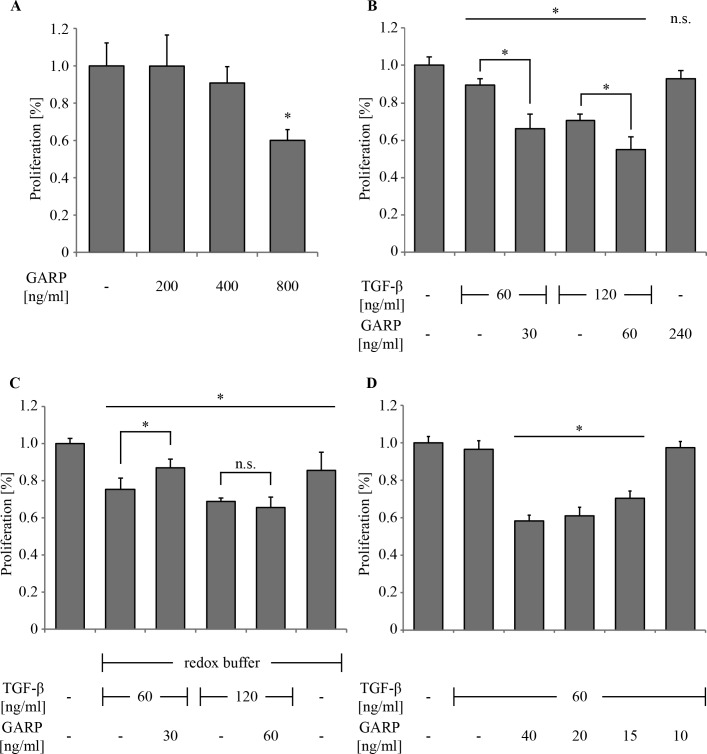
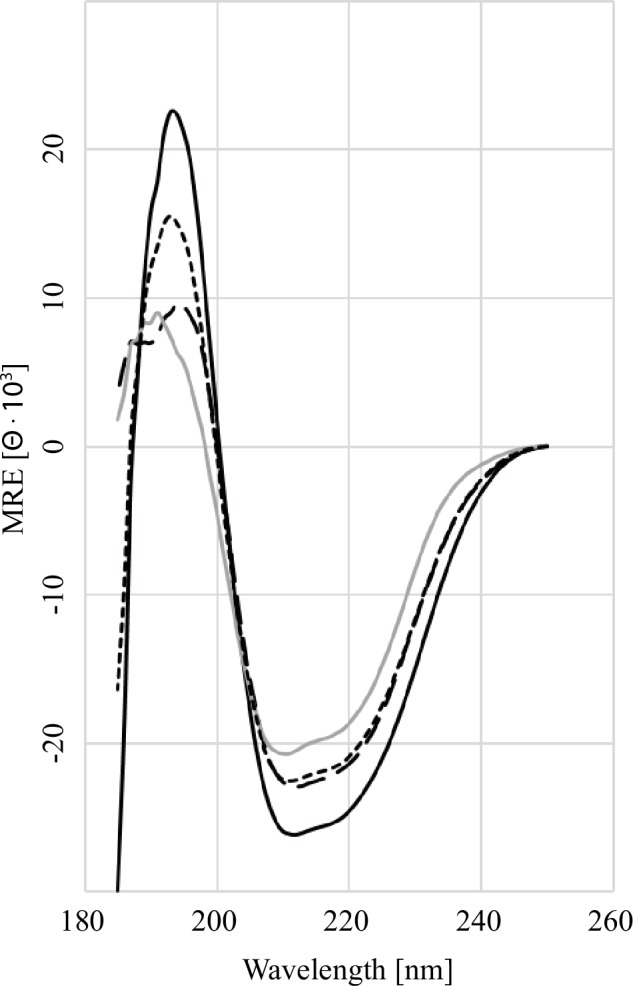
GARP (glycoprotein A repetitions predominant) is a cell surface receptor on regulatory T-lymphocytes, platelets, hepatic stellate cells and certain cancer cells. Its described function is the binding and accommodation of latent TGFβ (transforming growth factor), before the activation and release of the mature cytokine. For regulatory T cells it was shown that a knockdown of GARP or a treatment with blocking antibodies dramatically decreases their immune suppressive capacity. This confirms a fundamental role of GARP in the basic function of regulatory T cells. Prerequisites postulated for physiological GARP function include membrane anchorage of GARP, disulfide bridges between the propeptide of TGFβ and GARP and connection of this propeptide to αvβ6 or αvβ8 integrins of target cells during mechanical TGFβ release. Other studies indicate the existence of soluble GARP complexes and a functionality of soluble GARP alone. In order to clarify the underlying molecular mechanism, we expressed and purified recombinant TGFβ and a soluble variant of GARP. Surprisingly, soluble GARP and TGFβ formed stable non-covalent complexes in addition to disulfide-coupled complexes, depending on the redox conditions of the microenvironment. We also show that soluble GARP alone and the two variants of complexes mediate different levels of TGFβ activity. TGFβ activation is enhanced by the non-covalent GARP-TGFβ complex already at low (nanomolar) concentrations, at which GARP alone does not show any effect. This supports the idea of soluble GARP acting as immune modulator in vivo.
Conflict of interest statement
Figures
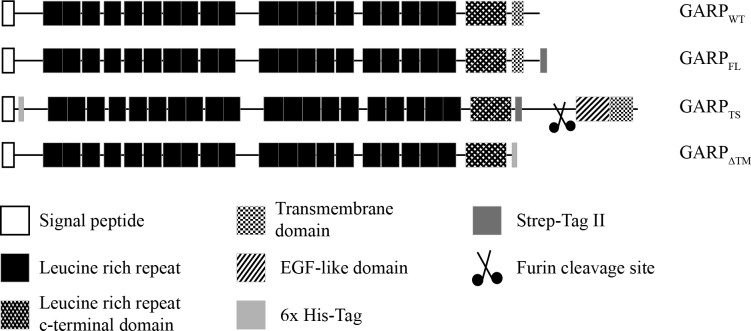
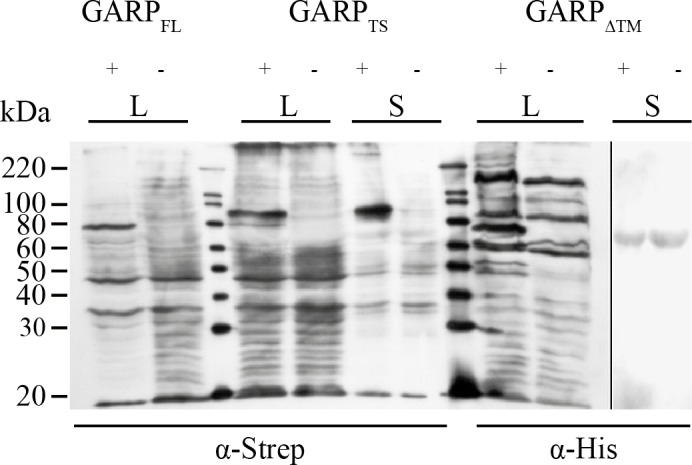
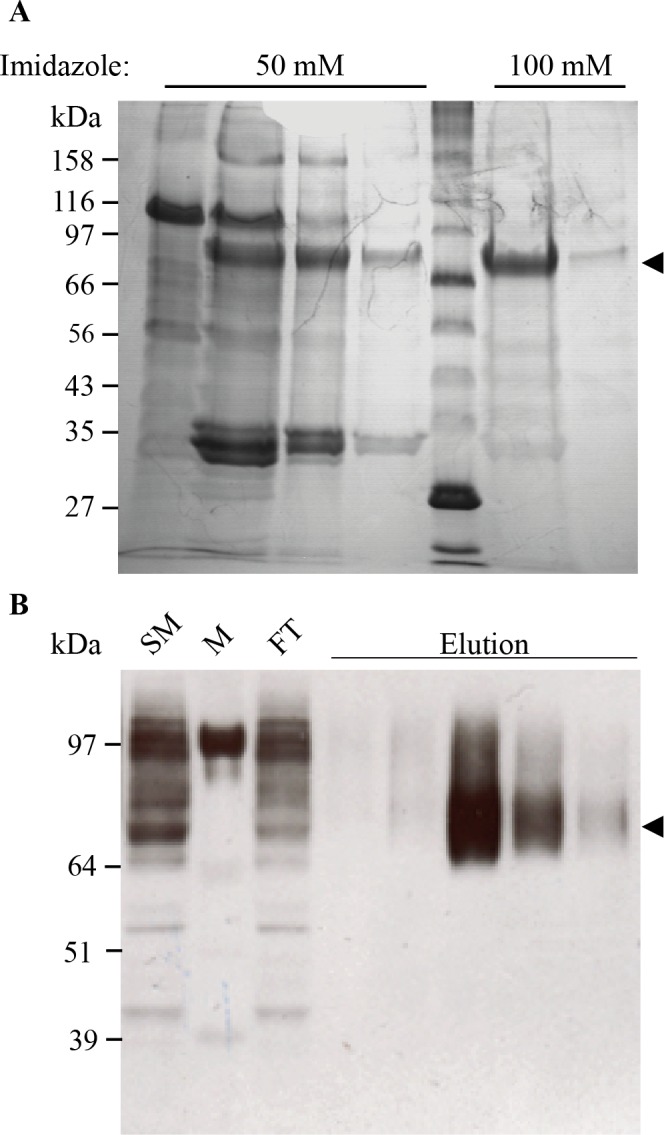
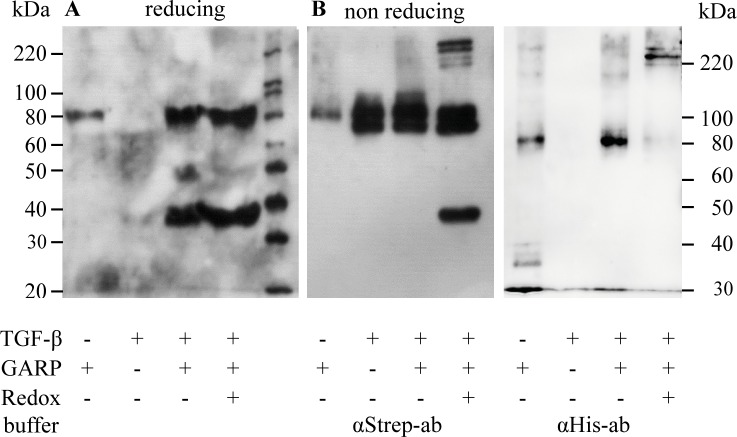
References
-
- Macaulay IC, Tijssen MR, Thijssen-timmer DC, Gusnanto A, Steward M, Burns P, et al. Comparative gene expression profiling of in vitro differentiated megakaryocytes and erythroblasts identifies novel activatory and inhibitory platelet membrane proteins. Blood. 2007; 3260–3269. 10.1182/blood-2006-07-036269 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
