

Regulatory T Cell Modulation by CBP/EP300 Bromodomain Inhibition
- PMID: 27056325
- PMCID: PMC4933219
- DOI: 10.1074/jbc.M115.708560
Regulatory T Cell Modulation by CBP/EP300 Bromodomain Inhibition
Abstract
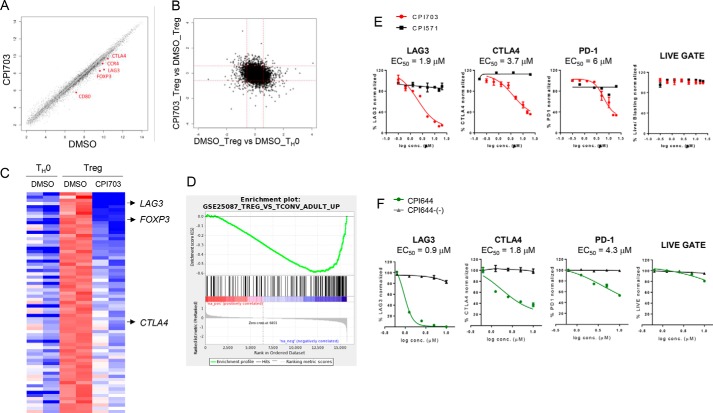
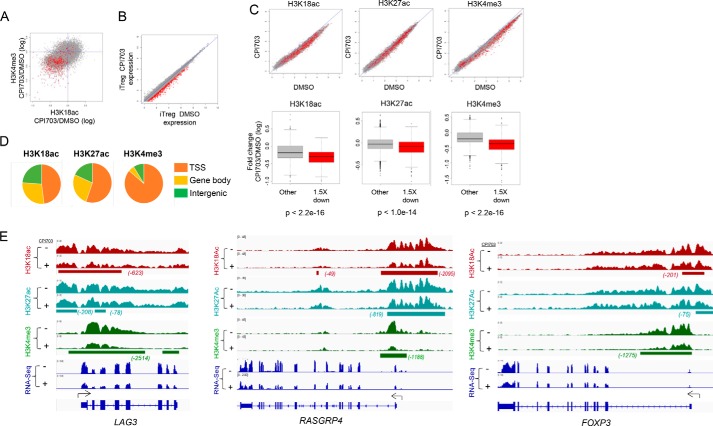
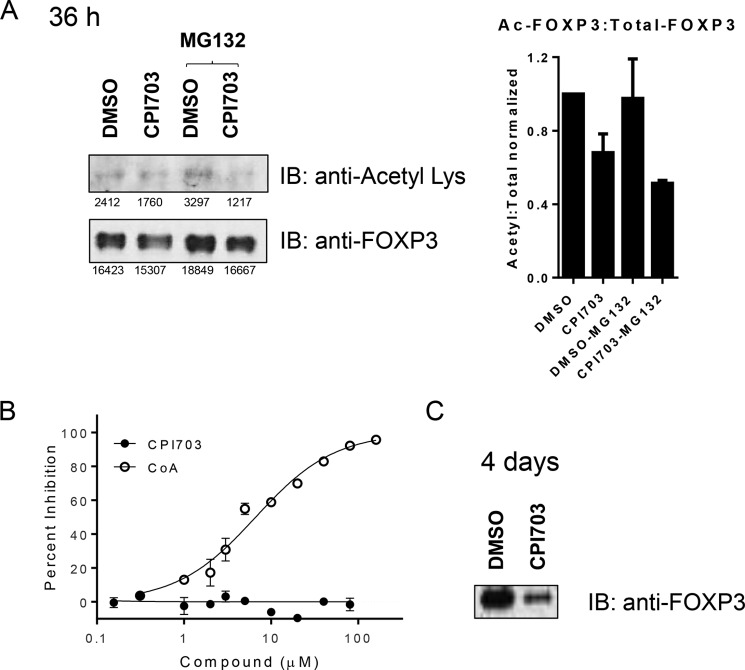
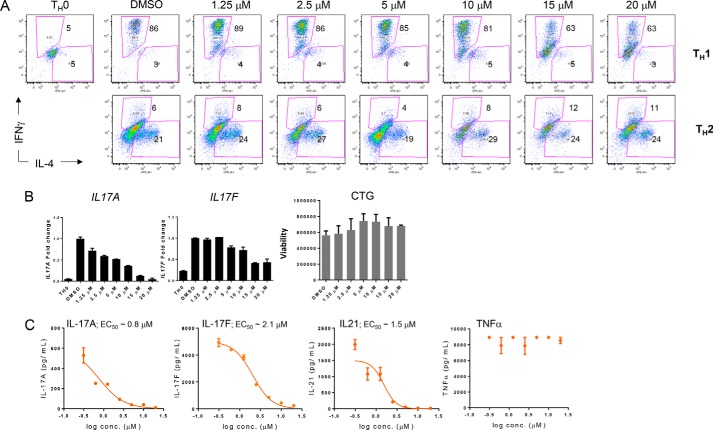
Covalent modification of histones is a fundamental mechanism of regulated gene expression in eukaryotes, and interpretation of histone modifications is an essential feature of epigenetic control. Bromodomains are specialized binding modules that interact with acetylated histones, linking chromatin recognition to gene transcription. Because of their ability to function in a domain-specific fashion, selective disruption of bromodomain:acetylated histone interactions with chemical probes serves as a powerful means for understanding biological processes regulated by these chromatin adaptors. Here we describe the discovery and characterization of potent and selective small molecule inhibitors for the bromodomains of CREBBP/EP300 that engage their target in cellular assays. We use these tools to demonstrate a critical role for CREBBP/EP300 bromodomains in regulatory T cell biology. Because regulatory T cell recruitment to tumors is a major mechanism of immune evasion by cancer cells, our data highlight the importance of CREBBP/EP300 bromodomain inhibition as a novel, small molecule-based approach for cancer immunotherapy.
Keywords: Treg, CBP, EP300, bromodomain, histone; acetylation; cellular immune response; chemical biology; chromatin regulation; forkhead box P3 (FOXP3).
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures
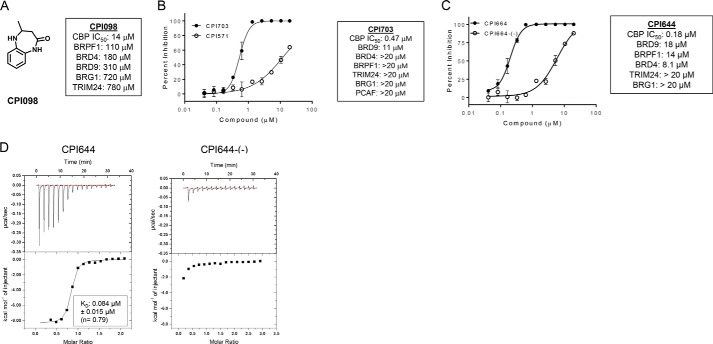
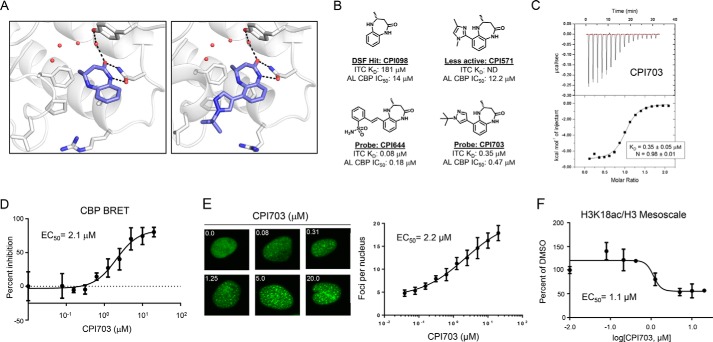
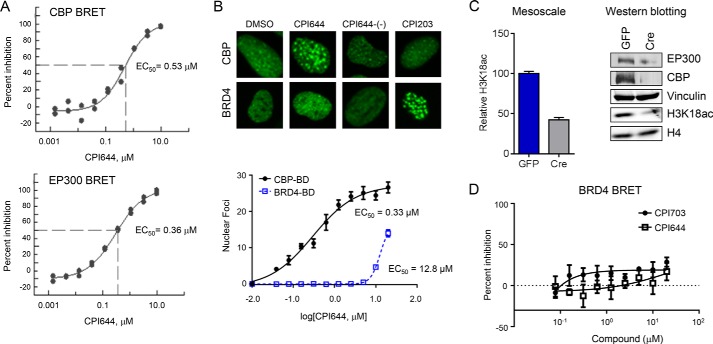
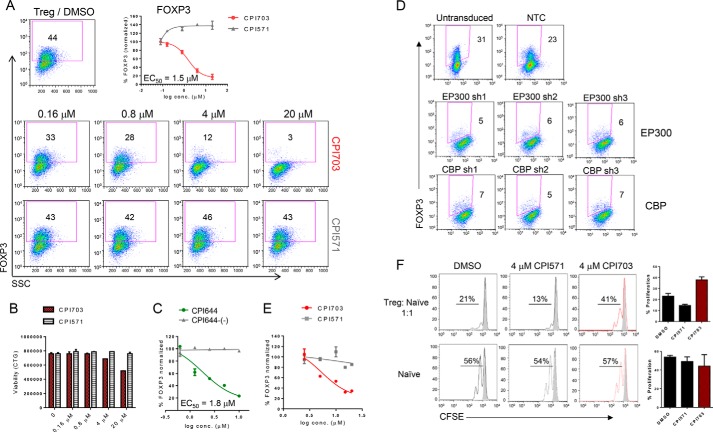
References
-
- Sakaguchi S., Ono M., Setoguchi R., Yagi H., Hori S., Fehervari Z., Shimizu J., Takahashi T., and Nomura T. (2006) Foxp3+ CD25+ CD4+ natural regulatory T cells in dominant self-tolerance and autoimmune disease. Immunol. Rev. 212, 8–27 - PubMed
-
- Zheng Y., and Rudensky A. Y. (2007) Foxp3 in control of the regulatory T cell lineage. Nat. Immunol. 8, 457–462 - PubMed
-
- Sugiyama D., Nishikawa H., Maeda Y., Nishioka M., Tanemura A., Katayama I., Ezoe S., Kanakura Y., Sato E., Fukumori Y., Karbach J., Jäger E., and Sakaguchi S. (2013) Anti-CCR4 mAb selectively depletes effector-type FoxP3+CD4+ regulatory T cells, evoking antitumor immune responses in humans. Proc. Natl. Acad. Sci. U.S.A. 110, 17945–17950 - PMC - PubMed
-
- Curiel T. J., Coukos G., Zou L., Alvarez X., Cheng P., Mottram P., Evdemon-Hogan M., Conejo-Garcia J. R., Zhang L., Burow M., Zhu Y., Wei S., Kryczek I., Daniel B., Gordon A., Myers L., Lackner A., Disis M. L., Knutson K. L., Chen L., and Zou W. (2004) Specific recruitment of regulatory T cells in ovarian carcinoma fosters immune privilege and predicts reduced survival. Nat. Med. 10, 942–949 - PubMed
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
