

Competition between Jagged-Notch and Endothelin1 Signaling Selectively Restricts Cartilage Formation in the Zebrafish Upper Face
- PMID: 27058748
- PMCID: PMC4825933
- DOI: 10.1371/journal.pgen.1005967
Competition between Jagged-Notch and Endothelin1 Signaling Selectively Restricts Cartilage Formation in the Zebrafish Upper Face
Abstract
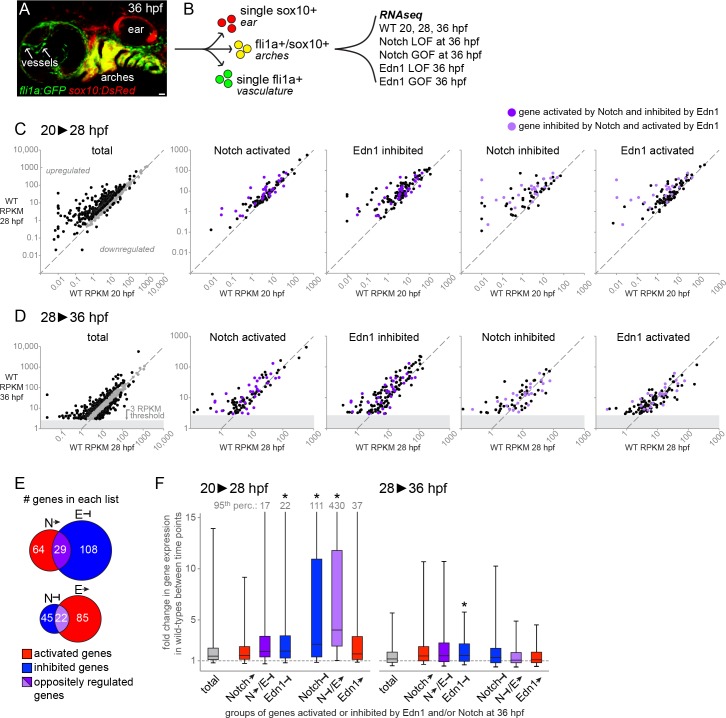
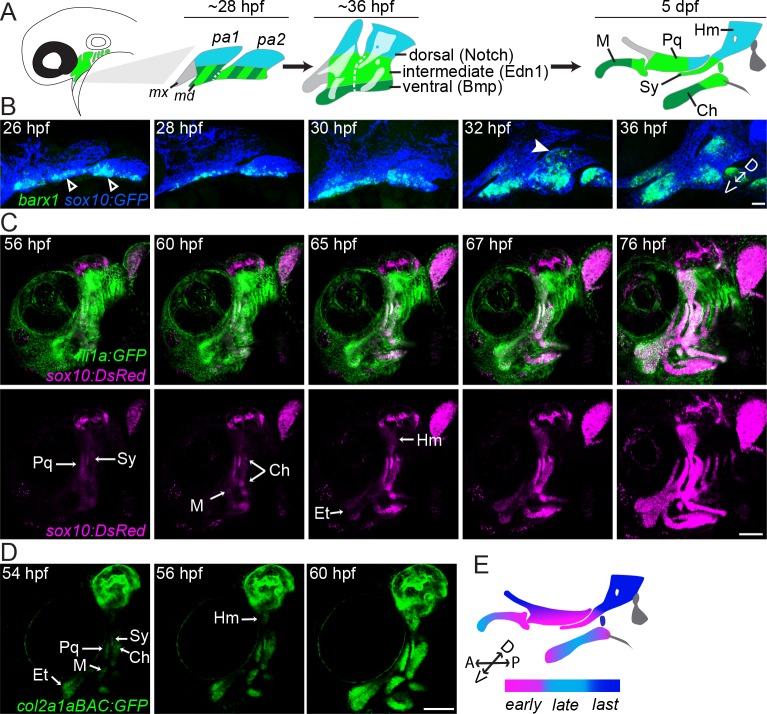
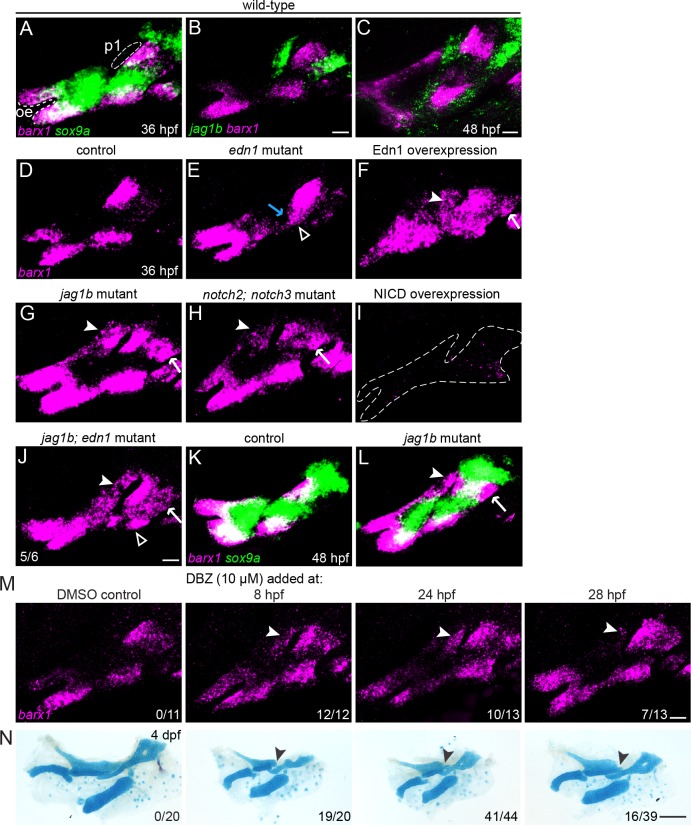
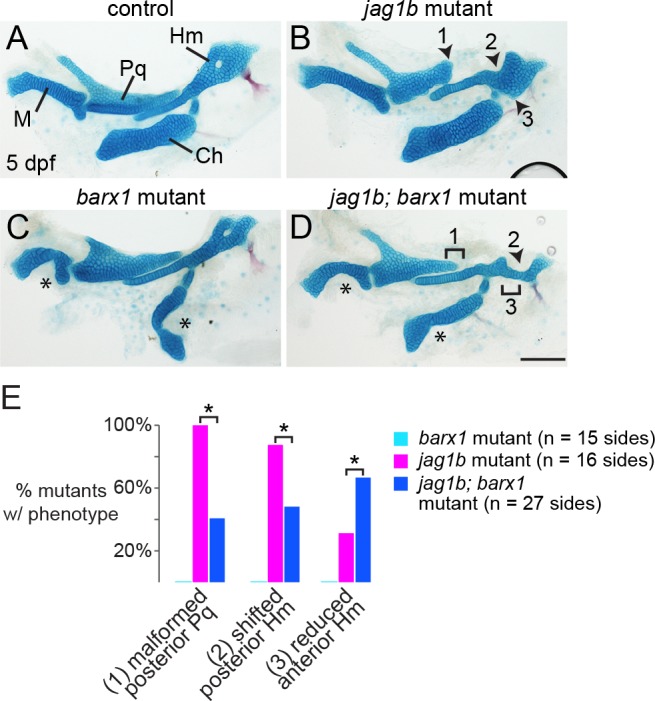
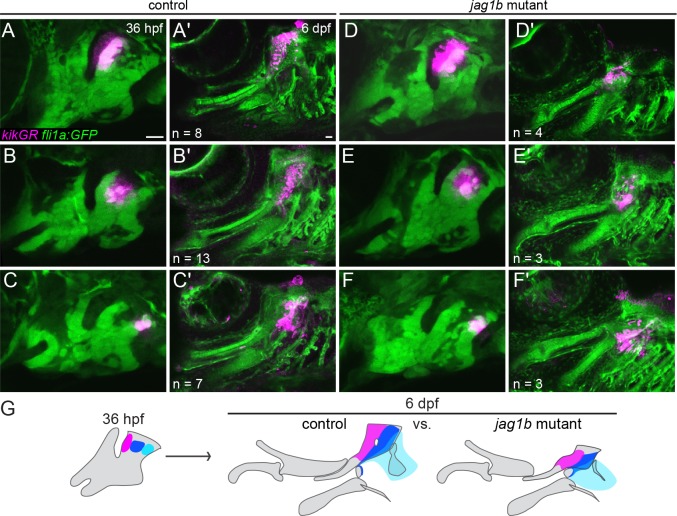
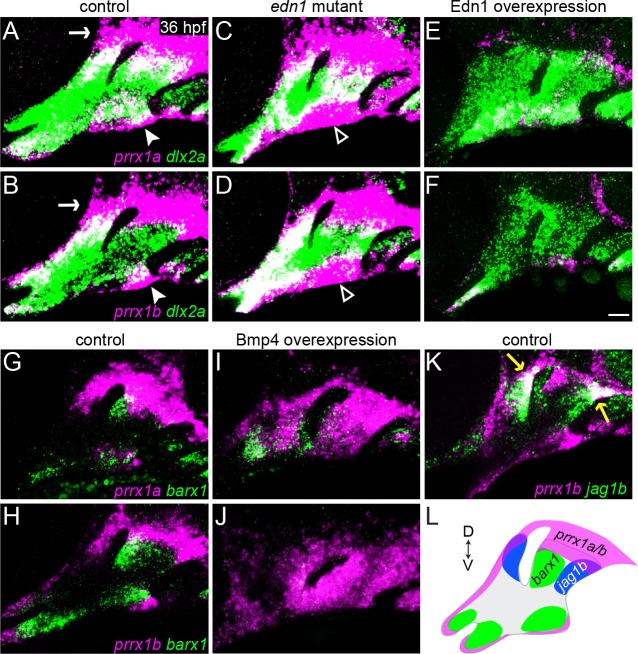
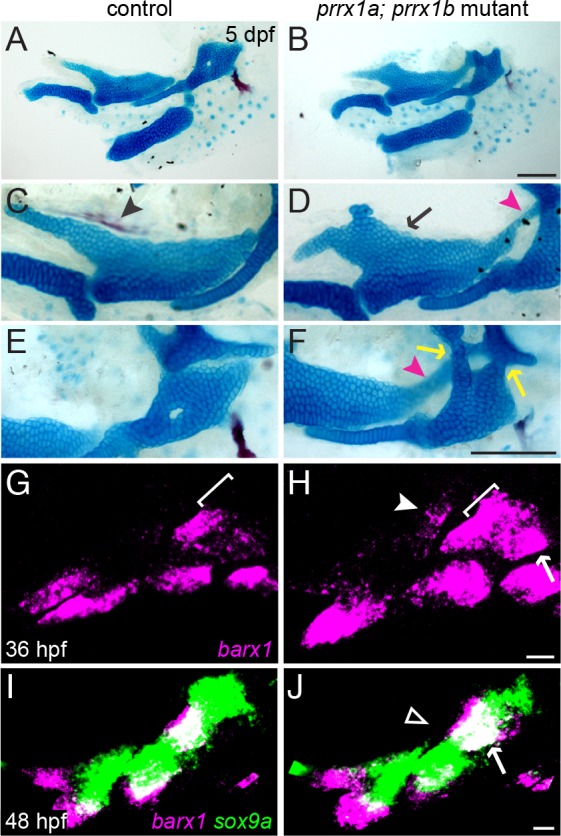
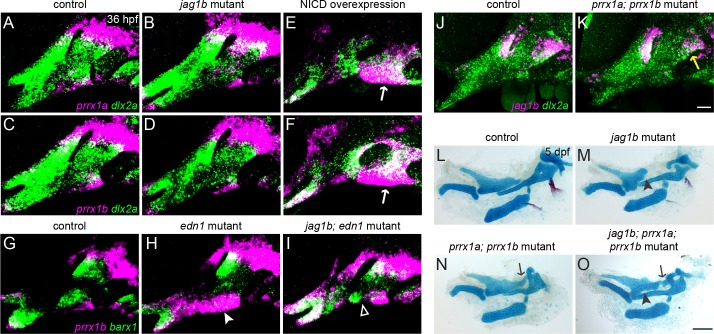
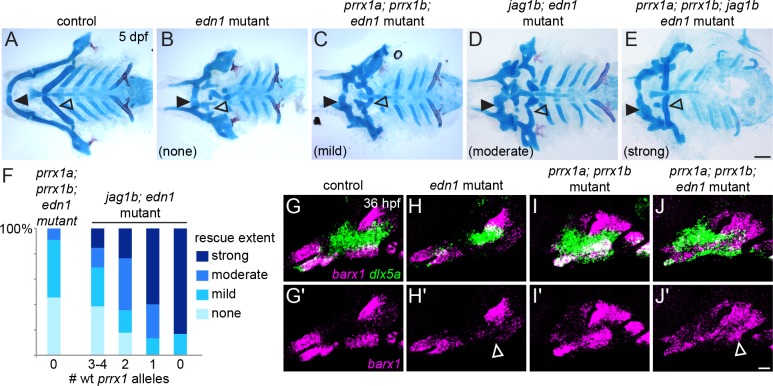
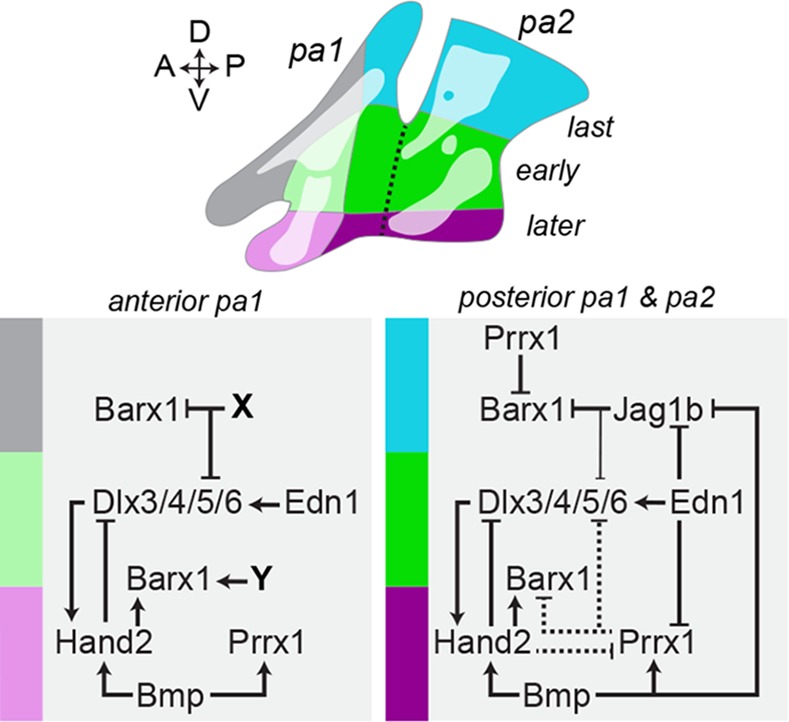
The intricate shaping of the facial skeleton is essential for function of the vertebrate jaw and middle ear. While much has been learned about the signaling pathways and transcription factors that control facial patterning, the downstream cellular mechanisms dictating skeletal shapes have remained unclear. Here we present genetic evidence in zebrafish that three major signaling pathways - Jagged-Notch, Endothelin1 (Edn1), and Bmp - regulate the pattern of facial cartilage and bone formation by controlling the timing of cartilage differentiation along the dorsoventral axis of the pharyngeal arches. A genomic analysis of purified facial skeletal precursors in mutant and overexpression embryos revealed a core set of differentiation genes that were commonly repressed by Jagged-Notch and induced by Edn1. Further analysis of the pre-cartilage condensation gene barx1, as well as in vivo imaging of cartilage differentiation, revealed that cartilage forms first in regions of high Edn1 and low Jagged-Notch activity. Consistent with a role of Jagged-Notch signaling in restricting cartilage differentiation, loss of Notch pathway components resulted in expanded barx1 expression in the dorsal arches, with mutation of barx1 rescuing some aspects of dorsal skeletal patterning in jag1b mutants. We also identified prrx1a and prrx1b as negative Edn1 and positive Bmp targets that function in parallel to Jagged-Notch signaling to restrict the formation of dorsal barx1+ pre-cartilage condensations. Simultaneous loss of jag1b and prrx1a/b better rescued lower facial defects of edn1 mutants than loss of either pathway alone, showing that combined overactivation of Jagged-Notch and Bmp/Prrx1 pathways contribute to the absence of cartilage differentiation in the edn1 mutant lower face. These findings support a model in which Notch-mediated restriction of cartilage differentiation, particularly in the second pharyngeal arch, helps to establish a distinct skeletal pattern in the upper face.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
-
- Clouthier DE, Hosoda K, Richardson JA, Williams SC, Yanagisawa H, Kuwaki T, et al. Cranial and cardiac neural crest defects in endothelin-A receptor-deficient mice. Development. 1998;125(5):813–24. Epub 1998/05/09. . - PubMed
-
- Yanagisawa H, Yanagisawa M, Kapur RP, Richardson JA, Williams SC, Clouthier DE, et al. Dual genetic pathways of endothelin-mediated intercellular signaling revealed by targeted disruption of endothelin converting enzyme-1 gene. Development. 1998;125(5):825–36. Epub 1998/05/09. . - PubMed
-
- Miller CT, Schilling TF, Lee K, Parker J, Kimmel CB. sucker encodes a zebrafish Endothelin-1 required for ventral pharyngeal arch development. Development. 2000;127(17):3815–28. Epub 2000/08/10. . - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
