

Cadmium and Secondary Structure-dependent Function of a Degron in the Pca1p Cadmium Exporter
- PMID: 27059957
- PMCID: PMC4933287
- DOI: 10.1074/jbc.M116.724930
Cadmium and Secondary Structure-dependent Function of a Degron in the Pca1p Cadmium Exporter
Abstract
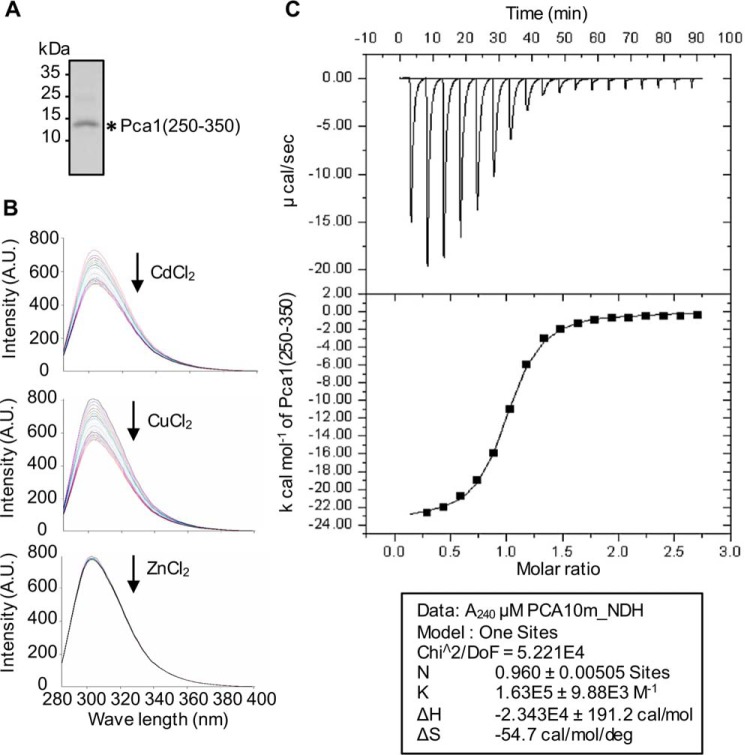
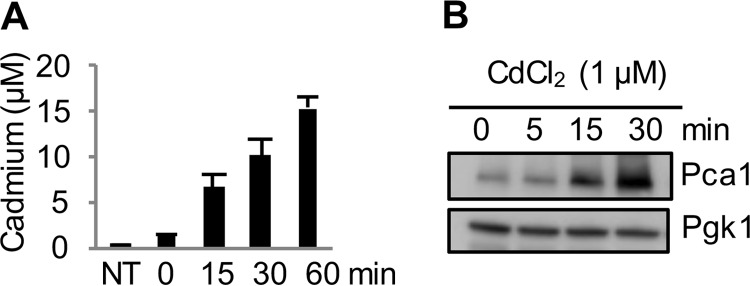
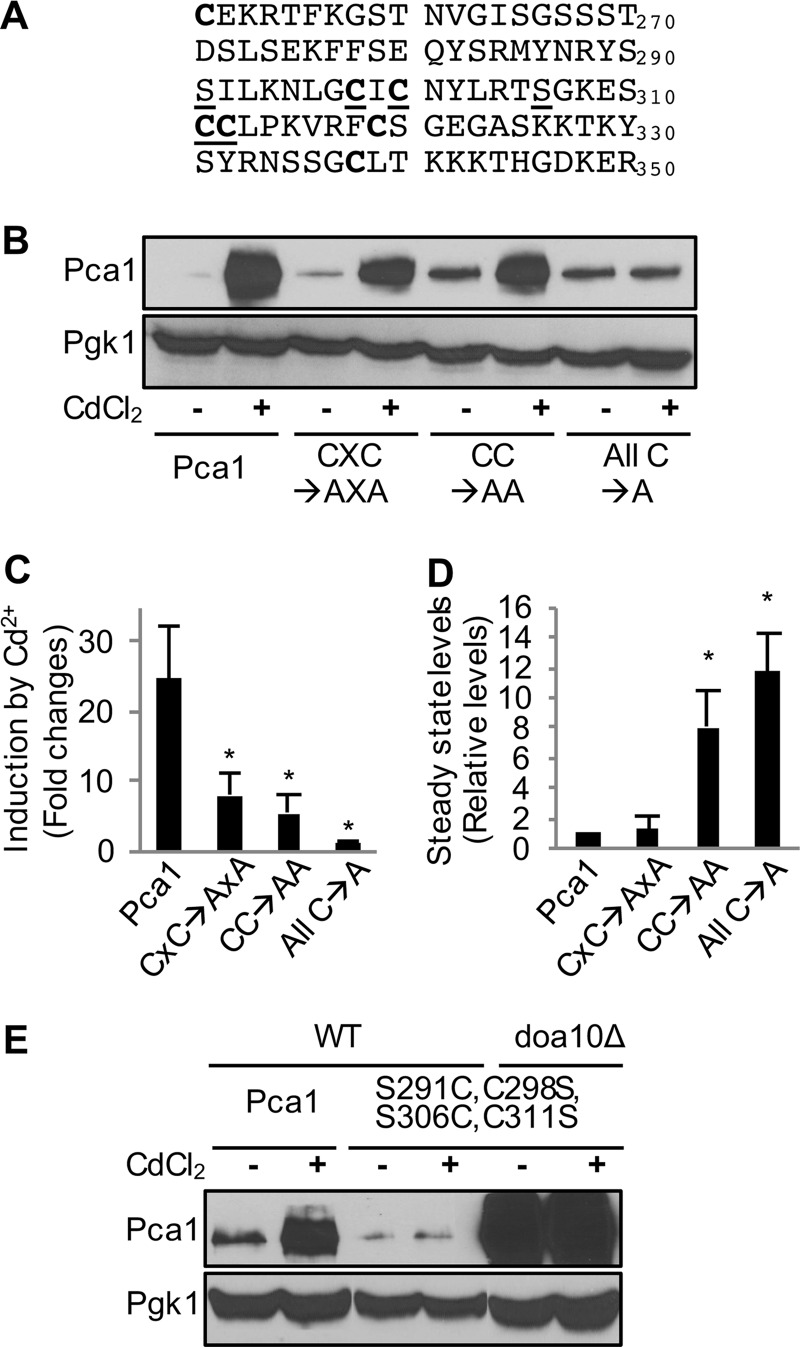
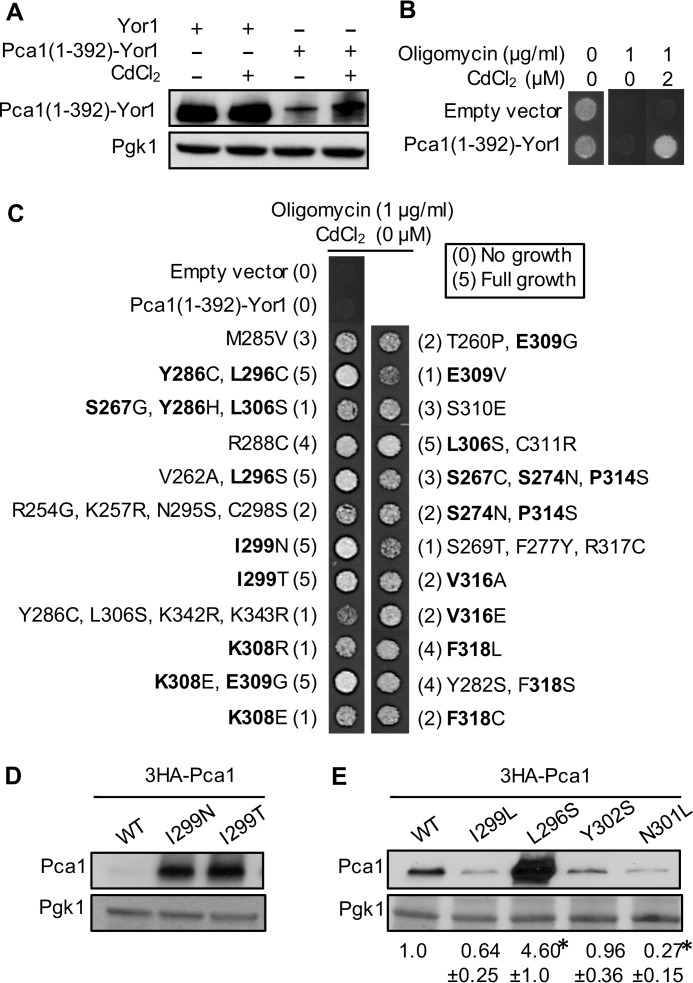
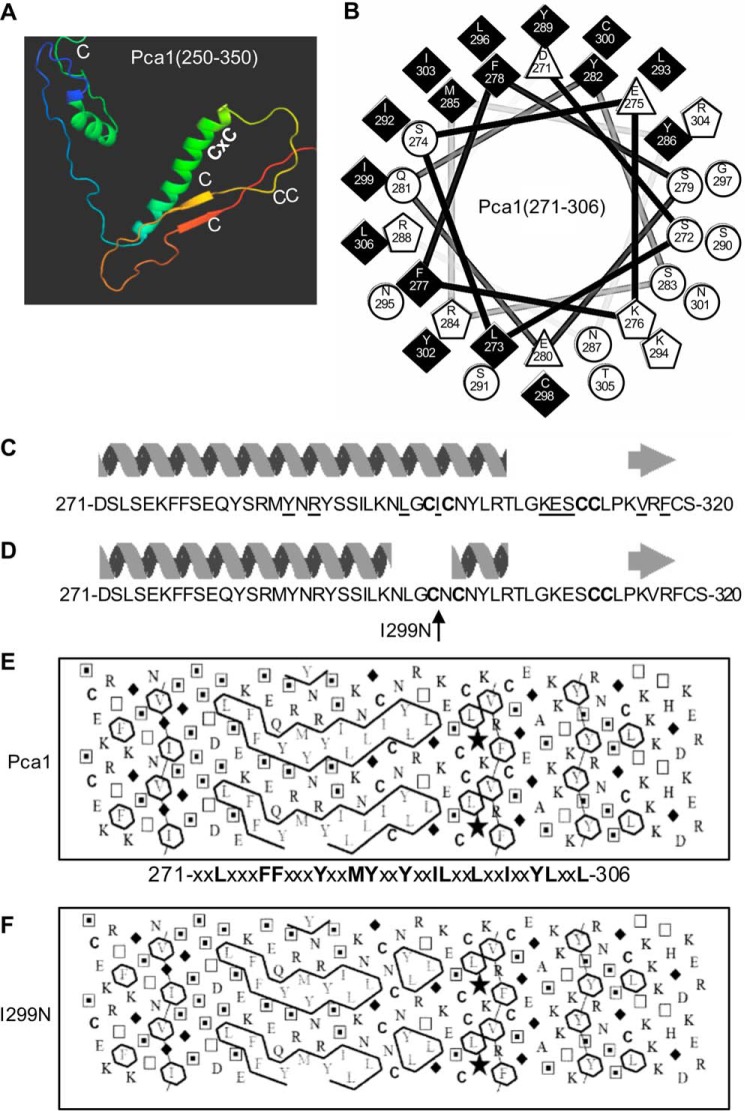
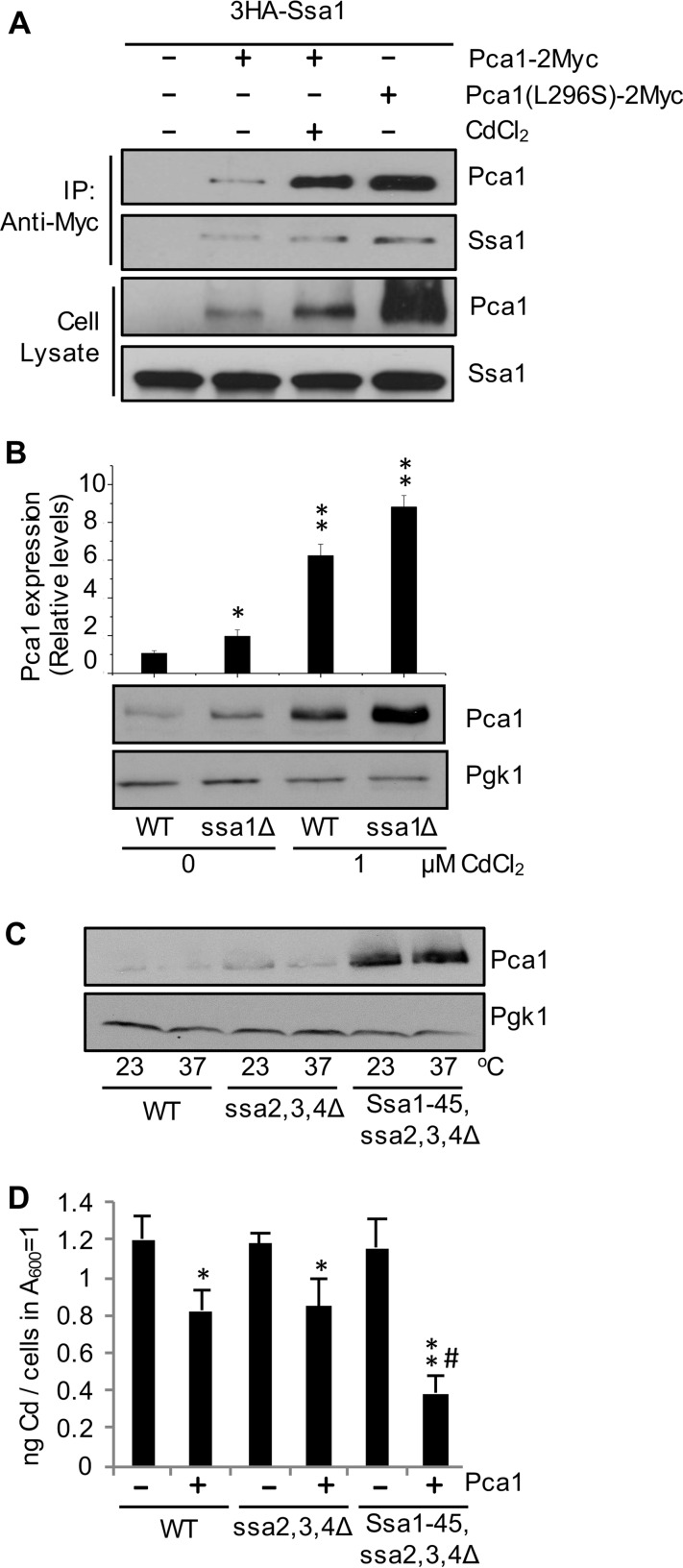
Protein turnover is a critical cellular process regulating biochemical pathways and destroying terminally misfolded or damaged proteins. Pca1p, a cadmium exporter in the yeast Saccharomyces cerevisiae, is rapidly degraded by the endoplasmic reticulum-associated degradation (ERAD) system via a cis-acting degron that exists at the 250-350 amino acid region of Pca1p and is transferable to other proteins to serve as a degradation signal. Cadmium stabilizes Pca1p in a manner dependent on the degron. This suggested that cadmium-mediated masking of the degron impedes its interaction with the molecular factors involved in the ERAD. The characteristics and mechanisms of action of the degron in Pca1p and most of those in other proteins however remain to be determined. The results presented here indicate that specific cysteine residues in a degron of Pca1p sense cadmium. An unbiased approach selecting non-functional degrons indicated a critical role of hydrophobic amino acids in the degron for its function. A secondary structure modeling predicted the formation of an amphipathic helix. Site-directed mutagenesis confirmed the functional significance of the hydrophobic patch. Last, hydrophobic amino acids in the degron- and cadmium-binding region affected the interaction of Pca1p with the Ssa1p molecular chaperone, which is involved in ERAD. These results reveal the mechanism of action of the degron, which might be useful for the identification and characterization of other degrons.
Keywords: ATPase; cadmium; degron; endoplasmic reticulum-associated protein degradation (ERAD); metal homeostasis; metal ion-protein interaction; molecular chaperone; transporter; yeast.
© 2016 by The American Society for Biochemistry and Molecular Biology, Inc.
Figures

Similar articles
-
Endoplasmic Reticulum-associated Degradation of Pca1p, a Polytopic Protein, via Interaction with the Proteasome at the Membrane.J Biol Chem. 2016 Jul 15;291(29):15082-92. doi: 10.1074/jbc.M116.726265. Epub 2016 May 12. J Biol Chem. 2016. PMID: 27226596 Free PMC article.
-
Cadmium-mediated rescue from ER-associated degradation induces expression of its exporter.Proc Natl Acad Sci U S A. 2009 Jun 23;106(25):10189-94. doi: 10.1073/pnas.0812114106. Epub 2009 Jun 10. Proc Natl Acad Sci U S A. 2009. PMID: 19515821 Free PMC article.
-
Degradation Signals for Ubiquitin-Proteasome Dependent Cytosolic Protein Quality Control (CytoQC) in Yeast.G3 (Bethesda). 2016 Jul 7;6(7):1853-66. doi: 10.1534/g3.116.027953. G3 (Bethesda). 2016. PMID: 27172186 Free PMC article.
-
Molecular chaperones HscA/Ssq1 and HscB/Jac1 and their roles in iron-sulfur protein maturation.Crit Rev Biochem Mol Biol. 2007 Mar-Apr;42(2):95-111. doi: 10.1080/10409230701322298. Crit Rev Biochem Mol Biol. 2007. PMID: 17453917 Review.
-
Atx1-like chaperones and their cognate P-type ATPases: copper-binding and transfer.Biometals. 2007 Jun;20(3-4):275-89. doi: 10.1007/s10534-006-9068-1. Epub 2007 Jan 16. Biometals. 2007. PMID: 17225061 Review.
Cited by
-
Effect of cadmium on essential metals and their impact on lipid metabolism in Saccharomyces cerevisiae.Cell Stress Chaperones. 2020 Jan;25(1):19-33. doi: 10.1007/s12192-019-01058-z. Epub 2019 Dec 10. Cell Stress Chaperones. 2020. PMID: 31823289 Free PMC article. Review.
-
Endoplasmic Reticulum-Associated Protein Degradation.Cold Spring Harb Perspect Biol. 2022 Dec 1;14(12):a041247. doi: 10.1101/cshperspect.a041247. Cold Spring Harb Perspect Biol. 2022. PMID: 35940909 Free PMC article. Review.
-
A conserved degron containing an amphipathic helix regulates the cholesterol-mediated turnover of human squalene monooxygenase, a rate-limiting enzyme in cholesterol synthesis.J Biol Chem. 2017 Dec 8;292(49):19959-19973. doi: 10.1074/jbc.M117.794230. Epub 2017 Sep 27. J Biol Chem. 2017. PMID: 28972164 Free PMC article.
-
The Hunt for Degrons of the 26S Proteasome.Biomolecules. 2019 Jun 13;9(6):230. doi: 10.3390/biom9060230. Biomolecules. 2019. PMID: 31200568 Free PMC article. Review.
-
Copper Resistance in Aspergillus nidulans Relies on the PI-Type ATPase CrpA, Regulated by the Transcription Factor AceA.Front Microbiol. 2017 May 30;8:912. doi: 10.3389/fmicb.2017.00912. eCollection 2017. Front Microbiol. 2017. PMID: 28611736 Free PMC article.
References
-
- Hochstrasser M. (1996) Protein degradation or regulation: Ub the judge. Cell 84, 813–815 - PubMed
-
- Zhang K., and Kaufman R. J. (2004) Signaling the unfolded protein response from the endoplasmic reticulum. J. Biol. Chem. 279, 25935–25938 - PubMed
-
- Ansari N., and Khodagholi F. (2013) Molecular mechanism aspect of ER stress in Alzheimer's disease: current approaches and future strategies. Curr. Drug Targets 14, 114–122 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
