

The Ku-binding motif is a conserved module for recruitment and stimulation of non-homologous end-joining proteins
- PMID: 27063109
- PMCID: PMC4831024
- DOI: 10.1038/ncomms11242
The Ku-binding motif is a conserved module for recruitment and stimulation of non-homologous end-joining proteins
Abstract
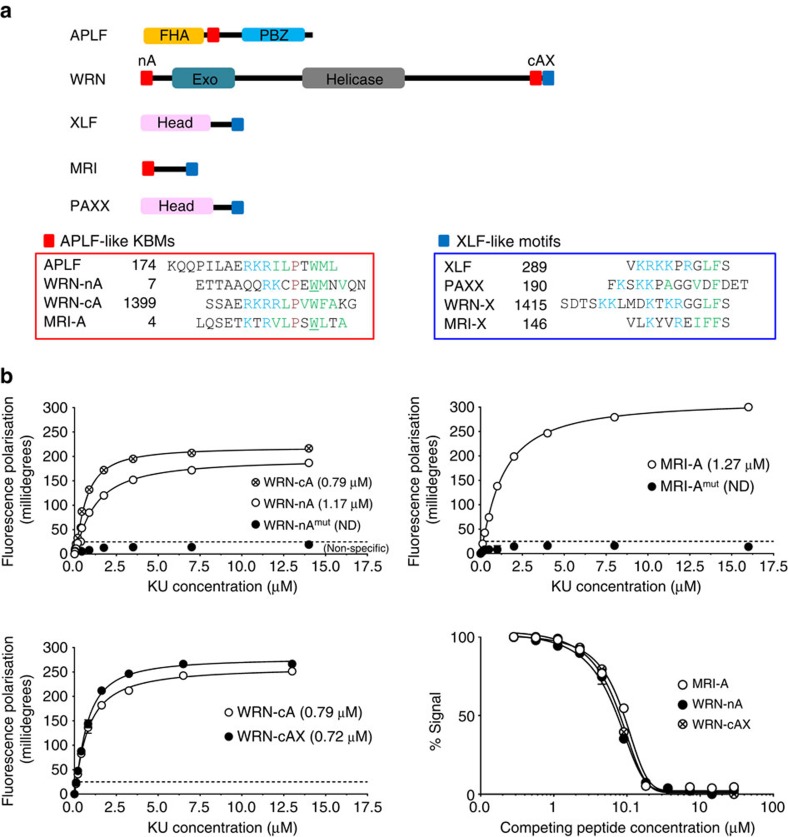
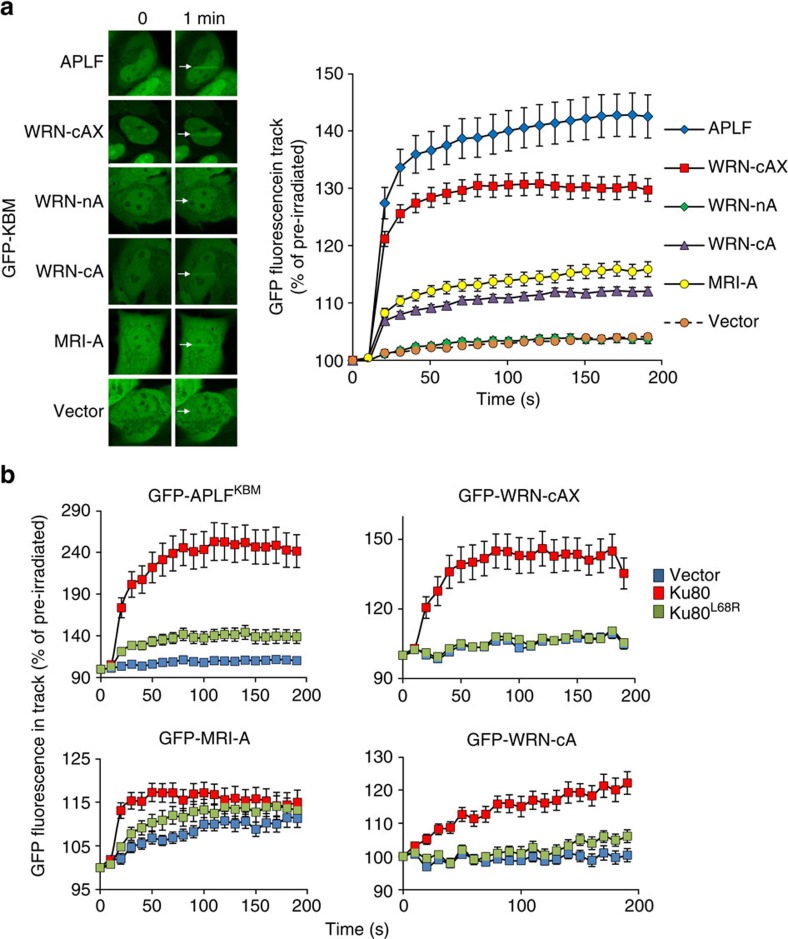
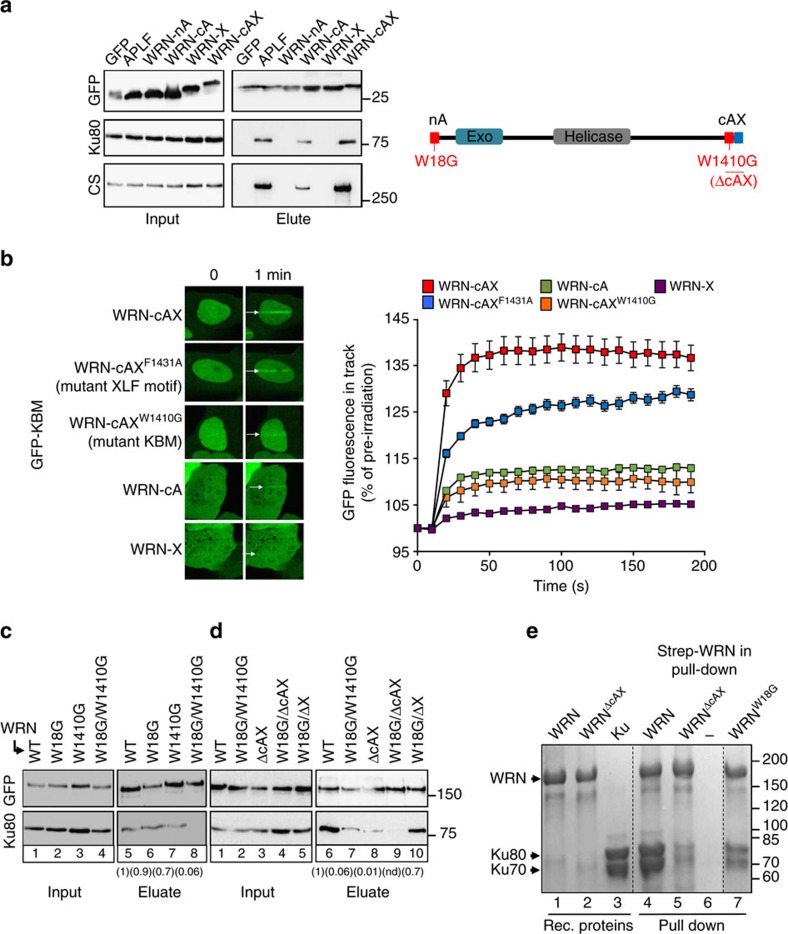
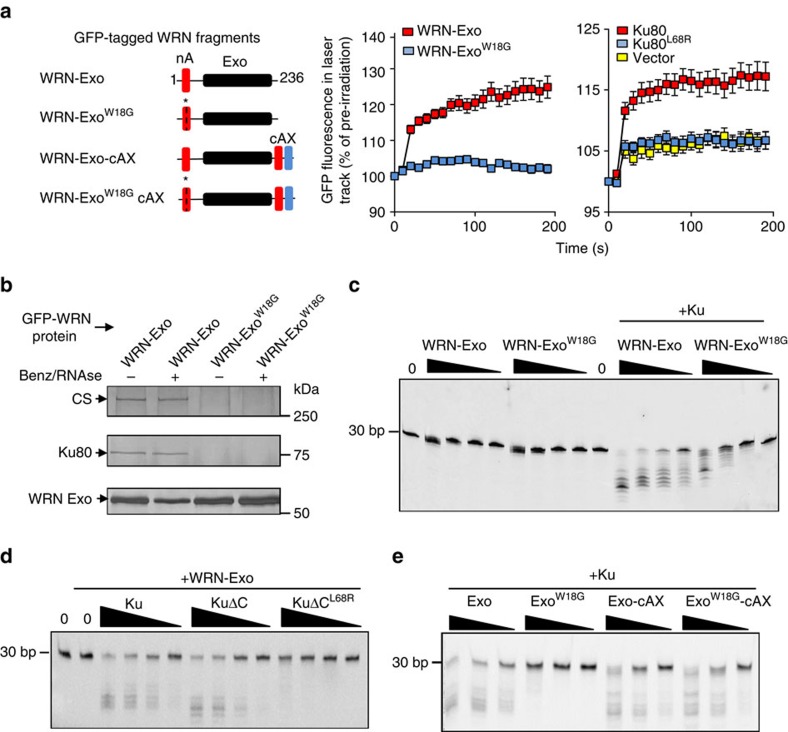
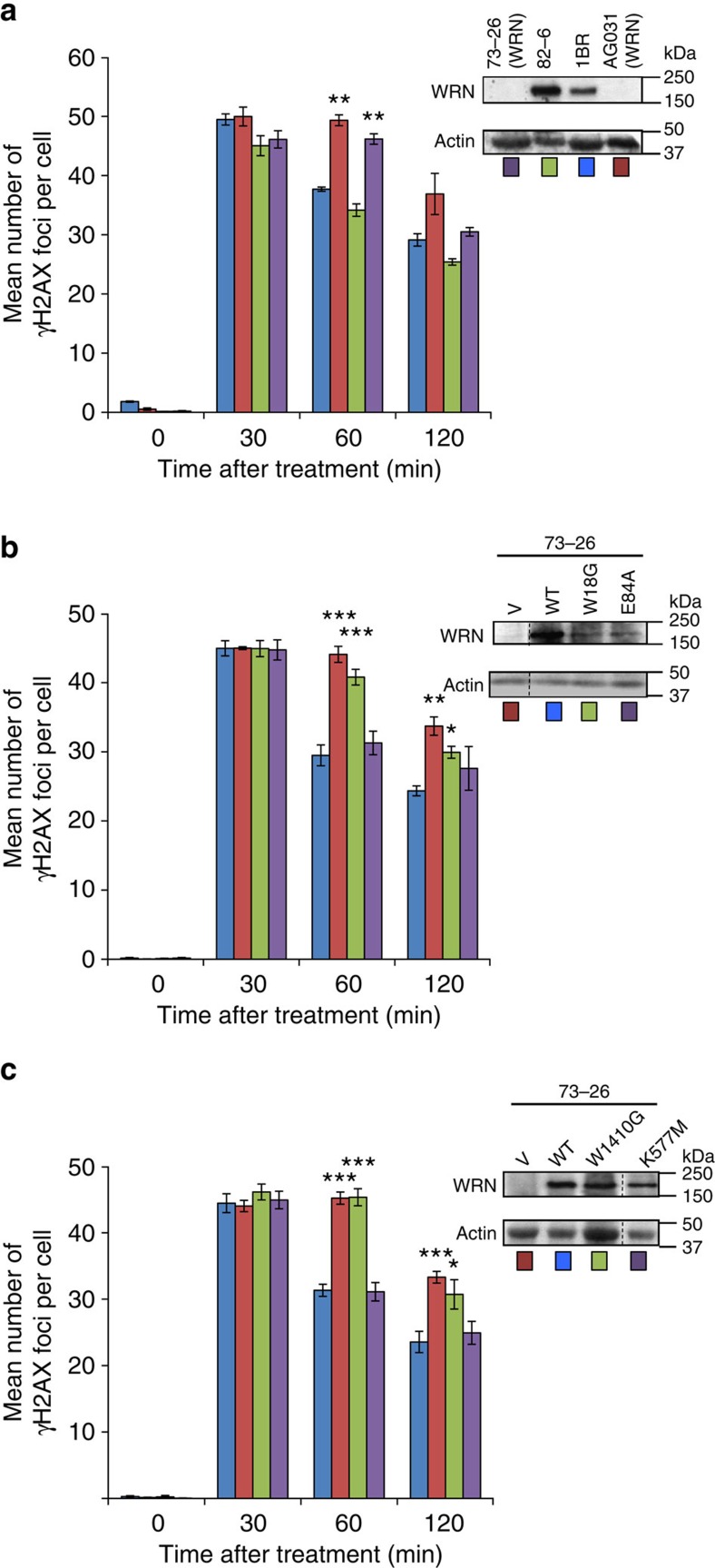
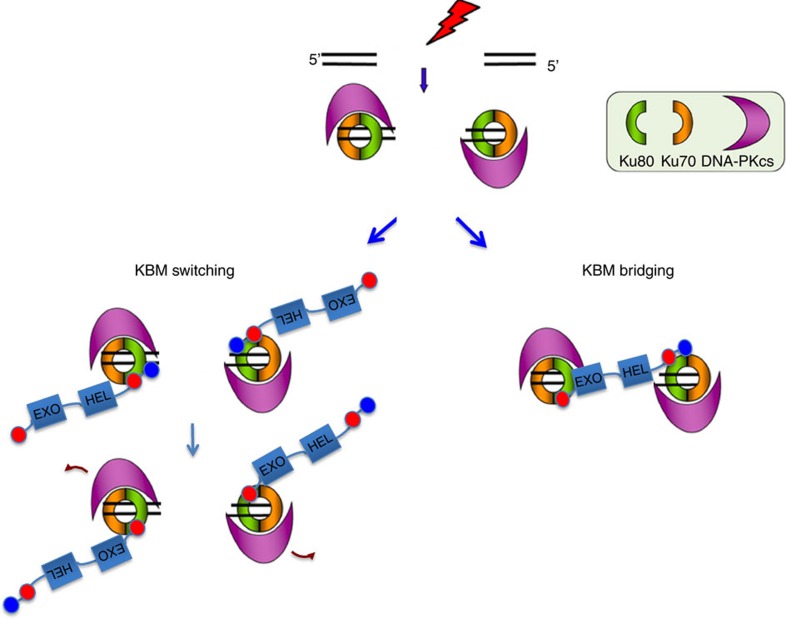
The Ku-binding motif (KBM) is a short peptide module first identified in APLF that we now show is also present in Werner syndrome protein (WRN) and in Modulator of retrovirus infection homologue (MRI). We also identify a related but functionally distinct motif in XLF, WRN, MRI and PAXX, which we denote the XLF-like motif. We show that WRN possesses two KBMs; one at the N terminus next to the exonuclease domain and one at the C terminus next to an XLF-like motif. We reveal that the WRN C-terminal KBM and XLF-like motif function cooperatively to bind Ku complexes and that the N-terminal KBM mediates Ku-dependent stimulation of WRN exonuclease activity. We also show that WRN accelerates DSB repair by a mechanism requiring both KBMs, demonstrating the importance of WRN interaction with Ku. These data define a conserved family of KBMs that function as molecular tethers to recruit and/or stimulate enzymes during NHEJ.
Figures
References
-
- Soulas-Sprauel P. et al. V(D)J and immunoglobulin class switch recombinations: a paradigm to study the regulation of DNA end-joining. Oncogene 26, 7780–7791 (2007). - PubMed
-
- McKinnon P. J. & Caldecott K. W. DNA strand break repair and human genetic disease. Annu. Rev. Genomics Hum. Genet. 8, 37–55 (2007). - PubMed
-
- Woodbine L., Gennery A. R. & Jeggo P. A. The clinical impact of deficiency in DNA non-homologous end-joining. DNA Repair (Amst) 16, 84–96 (2014). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
