

Unconventional endosome-like compartment and retromer complex in Toxoplasma gondii govern parasite integrity and host infection
- PMID: 27064065
- PMCID: PMC4831018
- DOI: 10.1038/ncomms11191
Unconventional endosome-like compartment and retromer complex in Toxoplasma gondii govern parasite integrity and host infection
Abstract
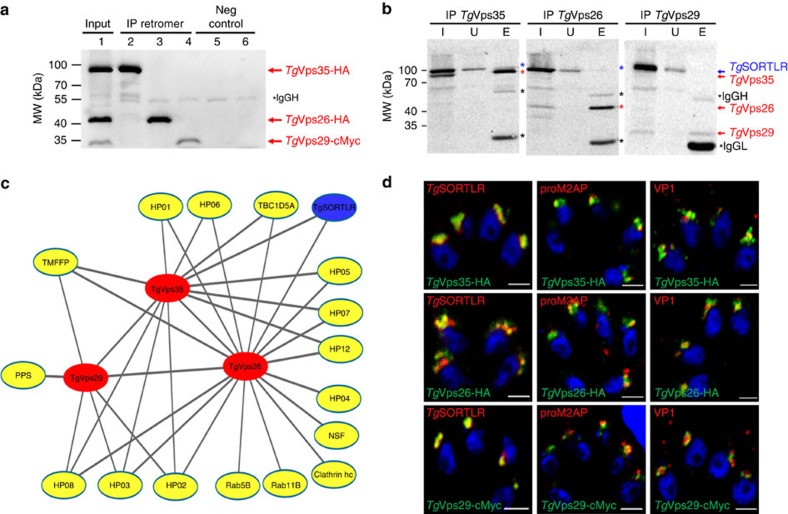
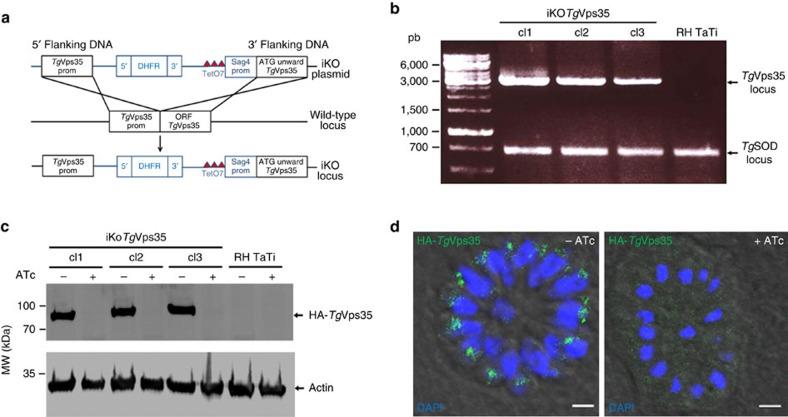
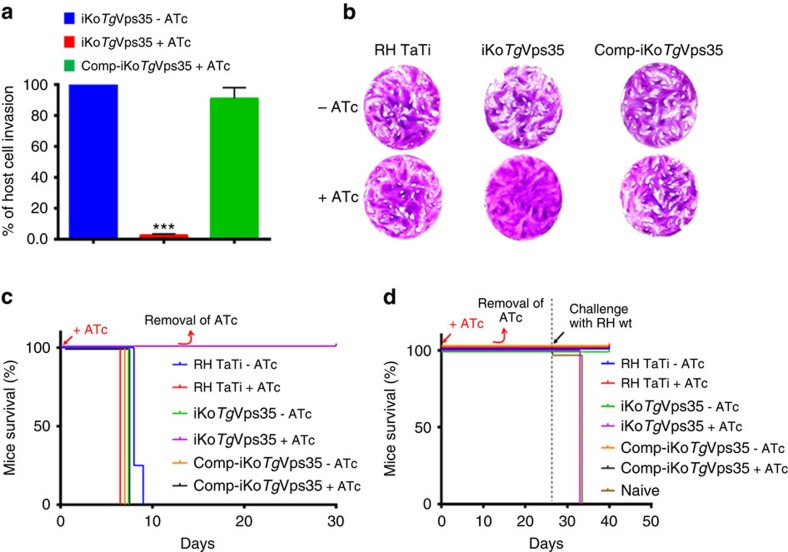
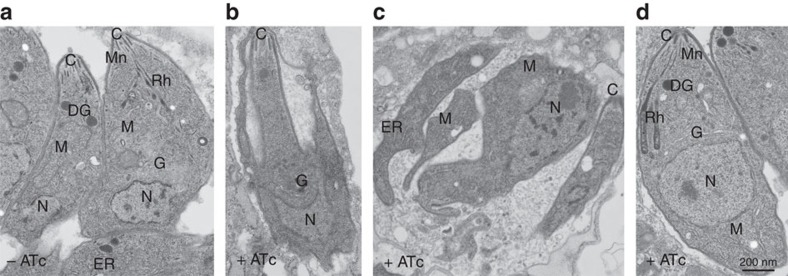
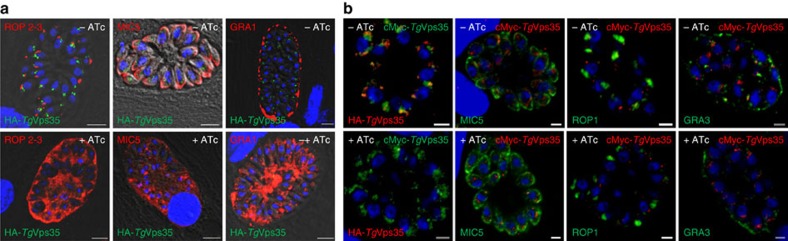
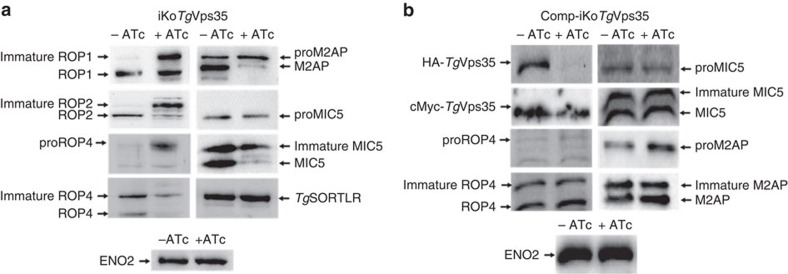
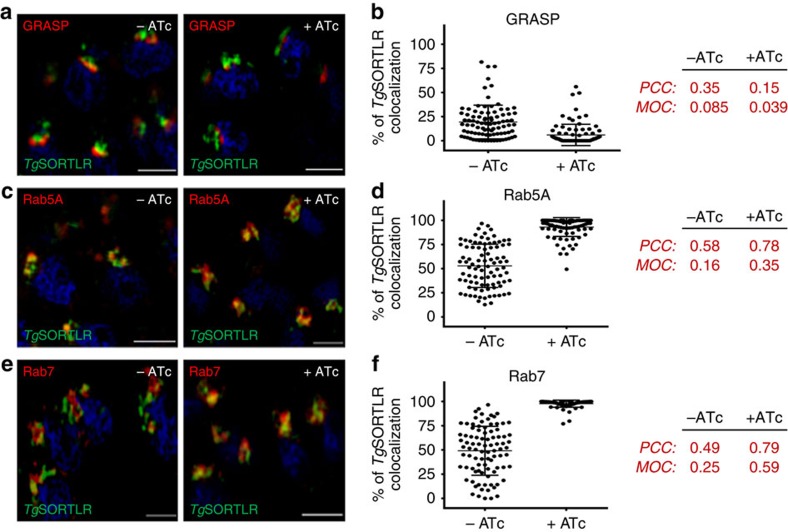
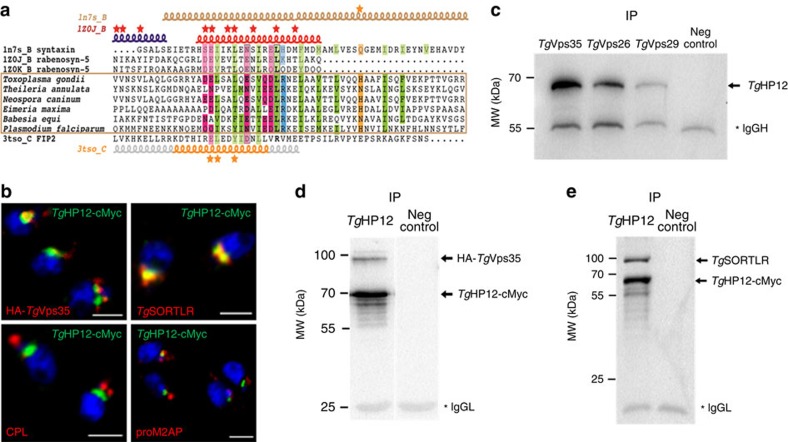
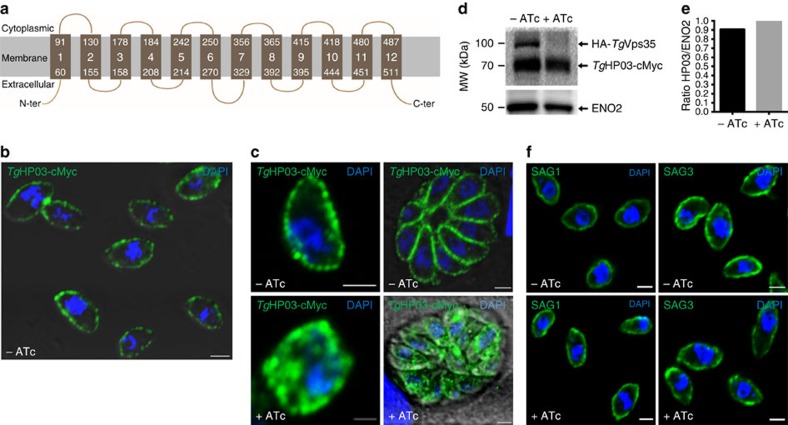
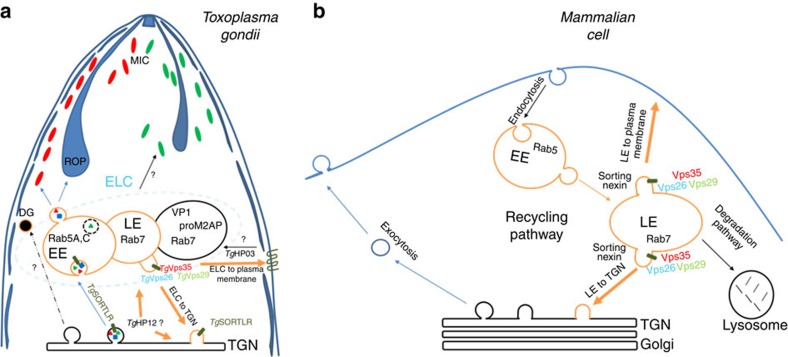
Membrane trafficking pathways play critical roles in Apicomplexa, a phylum of protozoan parasites that cause life-threatening diseases worldwide. Here we report the first retromer-trafficking interactome in Toxoplasma gondii. This retromer complex includes a trimer Vps35-Vps26-Vps29 core complex that serves as a hub for the endosome-like compartment and parasite-specific proteins. Conditional ablation of TgVps35 reveals that the retromer complex is crucial for the biogenesis of secretory organelles and for maintaining parasite morphology. We identify TgHP12 as a parasite-specific and retromer-associated protein with functions unrelated to secretory organelle formation. Furthermore, the major facilitator superfamily homologue named TgHP03, which is a multiple spanning and ligand transmembrane transporter, is maintained at the parasite membrane by retromer-mediated endocytic recycling. Thus, our findings highlight that both evolutionarily conserved and unconventional proteins act in concert in T. gondii by controlling retrograde transport that is essential for parasite integrity and host infection.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
Comment in
-
Toxoplasma Retromer Is Here to Stay.Trends Parasitol. 2016 Oct;32(10):758-760. doi: 10.1016/j.pt.2016.05.007. Epub 2016 May 27. Trends Parasitol. 2016. PMID: 27247246 Free PMC article.
References
-
- Pappas G., Roussos N. & Falagas M. E. Toxoplasmosis snapshots: Global status of Toxoplasma gondii seroprevalence and implications for pregnancy and congenital toxoplasmosis. Int. J. Parasitol. 39, 1385–1394 (2009). - PubMed
-
- Hager K. M., Striepen B., Tilney L. G. & Roos D. S. The nuclear envelope serves as an intermediary between the ER and Golgi complex in the intracellular parasite Toxoplasma gondii. J. Cell Sci. 112, 2631–2638 (1999). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
