

Nod2-mediated recognition of the microbiota is critical for mucosal adjuvant activity of cholera toxin
- PMID: 27064448
- PMCID: PMC4860092
- DOI: 10.1038/nm.4075
Nod2-mediated recognition of the microbiota is critical for mucosal adjuvant activity of cholera toxin
Erratum in
-
Corrigendum: Nod2-mediated recognition of the microbiota is critical for mucosal adjuvant activity of cholera toxin.Nat Med. 2016 Aug 4;22(8):961. doi: 10.1038/nm0816-961. Nat Med. 2016. PMID: 27490437 No abstract available.
Abstract
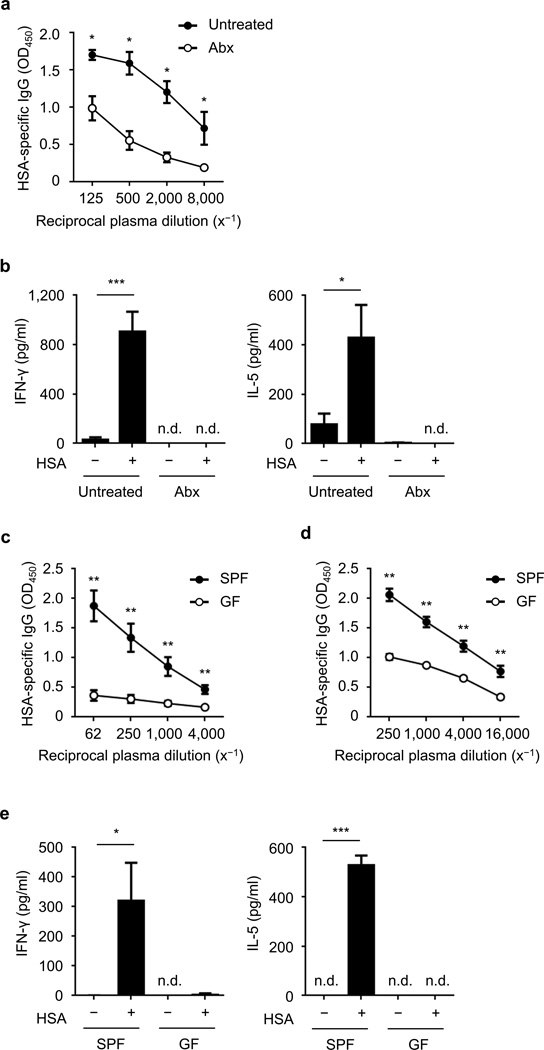
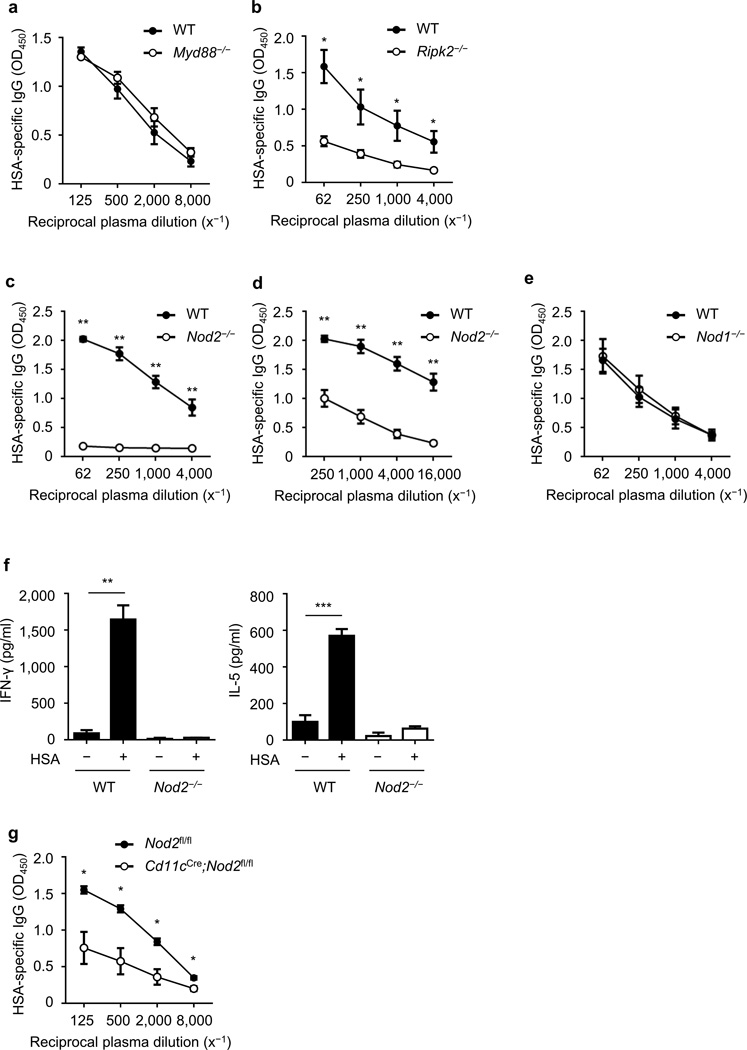
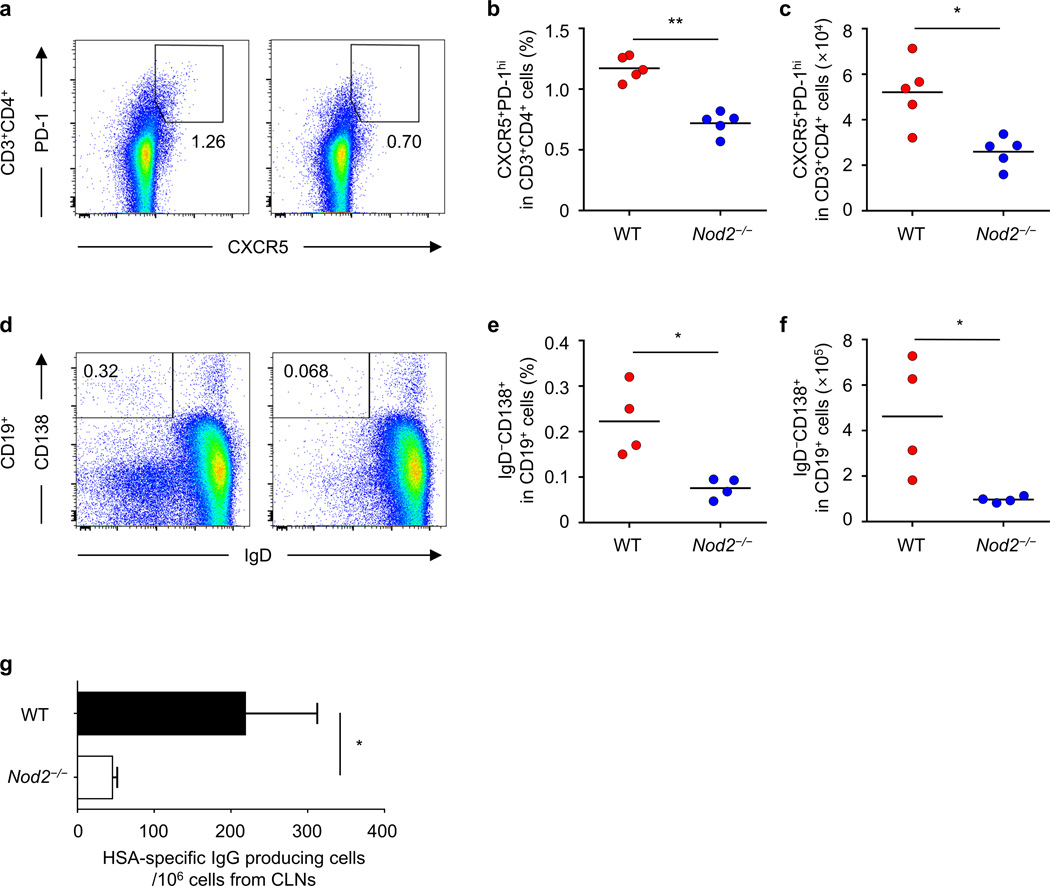
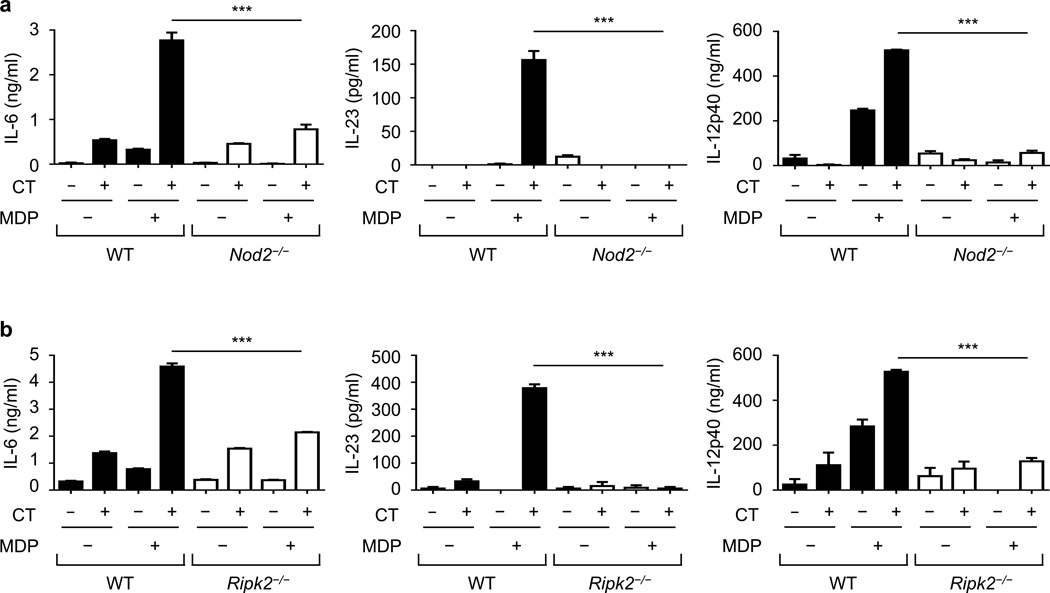
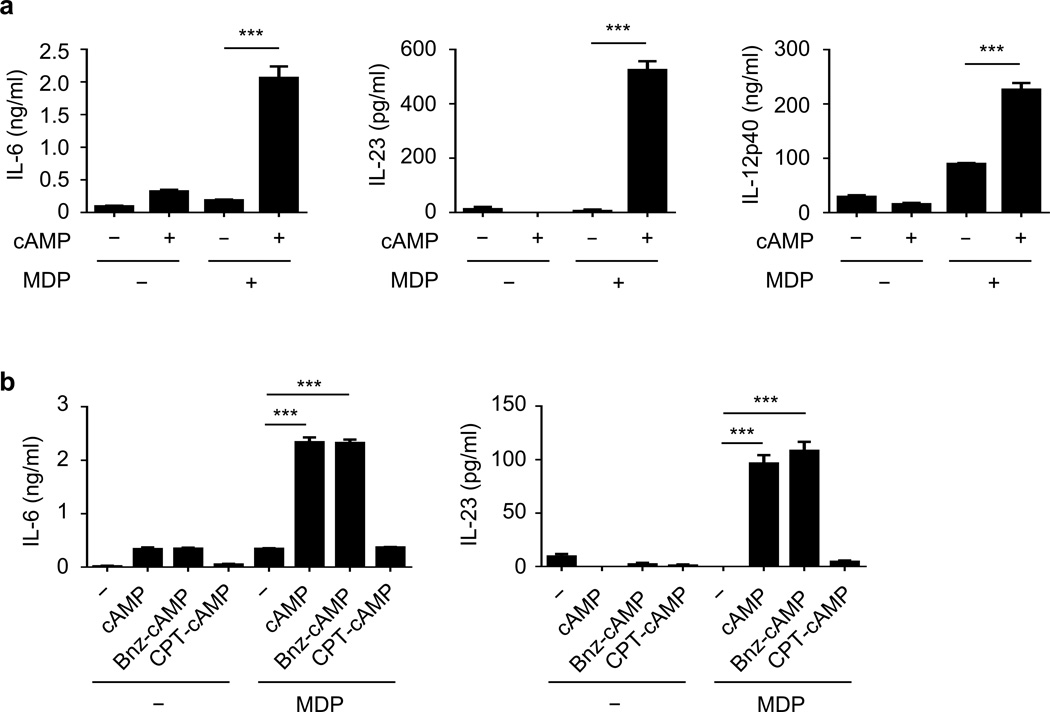
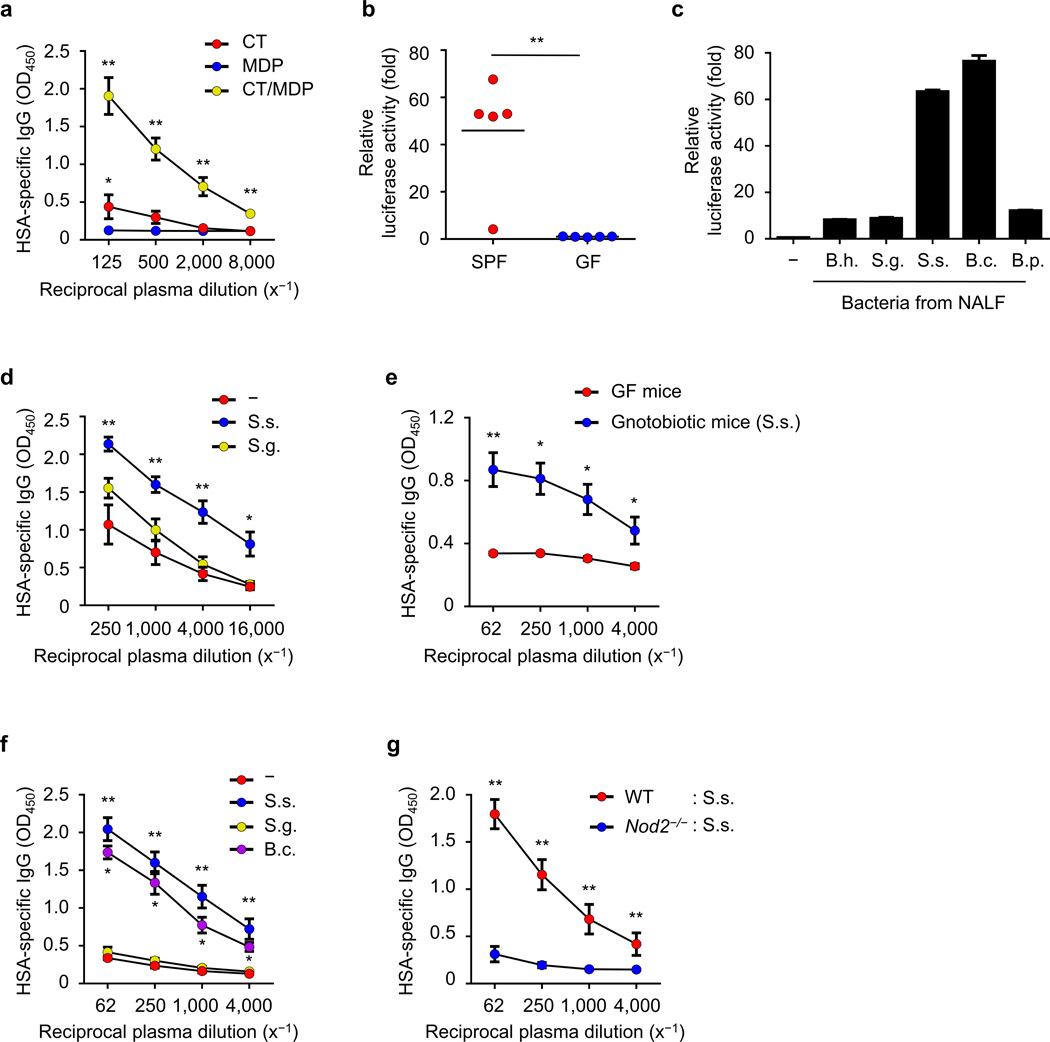
Cholera toxin (CT) is a potent adjuvant for inducing mucosal immune responses. However, the mechanism by which CT induces adjuvant activity remains unclear. Here we show that the microbiota is critical for inducing antigen-specific IgG production after intranasal immunization. After mucosal vaccination with CT, both antibiotic-treated and germ-free (GF) mice had reduced amounts of antigen-specific IgG, smaller recall-stimulated cytokine responses, impaired follicular helper T (TFH) cell responses and reduced numbers of plasma cells. Recognition of symbiotic bacteria via the nucleotide-binding oligomerization domain containing 2 (Nod2) sensor in cells that express the integrin CD11c (encoded by Itgax) was required for the adjuvanticity of CT. Reconstitution of GF mice with a Nod2 agonist or monocolonization with Staphylococcus sciuri, which has high Nod2-stimulatory activity, was sufficient to promote robust CT adjuvant activity, whereas bacteria with low Nod2-stimulatory activity did not. Mechanistically, CT enhanced Nod2-mediated cytokine production in dendritic cells via intracellular cyclic AMP. These results show a role for the microbiota and the intracellular receptor Nod2 in promoting the mucosal adjuvant activity of CT.
Figures
References
-
- Freytag LC, Clements JD. Mucosal adjuvants. Vaccine. 2005;23:1804–1813. - PubMed
-
- Elson CO, Ealding W. Generalized systemic and mucosal immunity in mice after mucosal stimulation with cholera toxin. J. Immunol. 1984;132:2736–2741. - PubMed
-
- Porgador A, Staats HF, Itoh Y, Kelsall BL. Intranasal immunization with cytotoxic T-lymphocyte epitope peptide and mucosal adjuvant cholera toxin: selective augmentation of peptide-presenting dendritic cells in nasal mucosa-associated lymphoid tissue. Infect. Immun. 1998;66:5876–5881. - PMC - PubMed
-
- Sunahara RK, Dessauer CW, Whisnant RE, Kleuss C, Gilman AG. Interaction of Gs with the Cytosolic Domains of Mammalian Adenylyl Cyclase. J. Biol. Chem. 1997;272:22265–22271. - PubMed
-
- Lycke N, Tsuji T, Holmgren J. The adjuvant effect of Vibrio cholerae and Escherichia coli heat-labile enterotoxins is linked to their ADP-ribosyltransferase activity. Eur. J. Immunol. 1992;22:2277–2281. - PubMed
Methods References
-
- Kim D, et al. Suppression of Allergic Diarrhea in Murine Ovalbumin-Induced Allergic Diarrhea Model by PG102, a Water-Soluble Extract Prepared from Actinidia arguta. Int. Arch. Allergy Immunol. 2009;150:164–171. - PubMed
-
- Wu HY, Nguyen HH, Russell MW. Nasal lymphoid tissue (NALT) as a mucosal immune inductive site. Scandinavian journal of immunology. 1997;46:506–513. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
