

Type III Interferons Produced by Human Placental Trophoblasts Confer Protection against Zika Virus Infection
- PMID: 27066743
- PMCID: PMC4866896
- DOI: 10.1016/j.chom.2016.03.008
Type III Interferons Produced by Human Placental Trophoblasts Confer Protection against Zika Virus Infection
Abstract
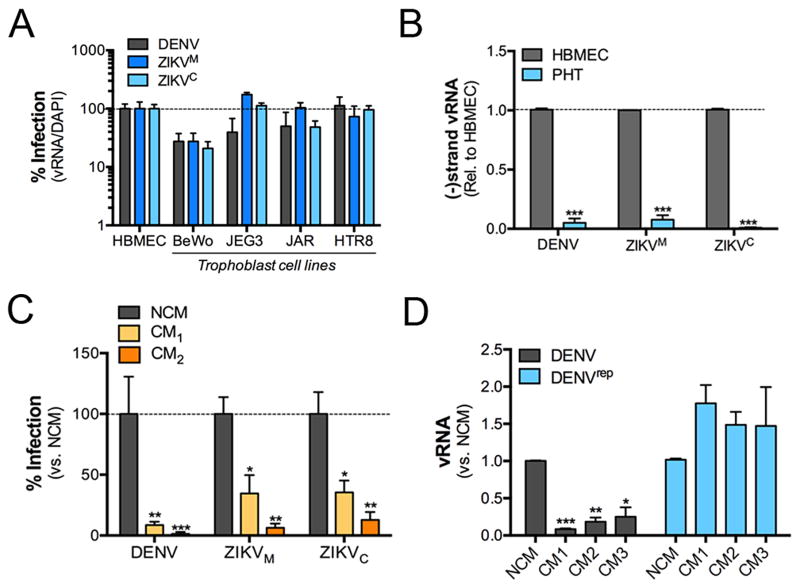
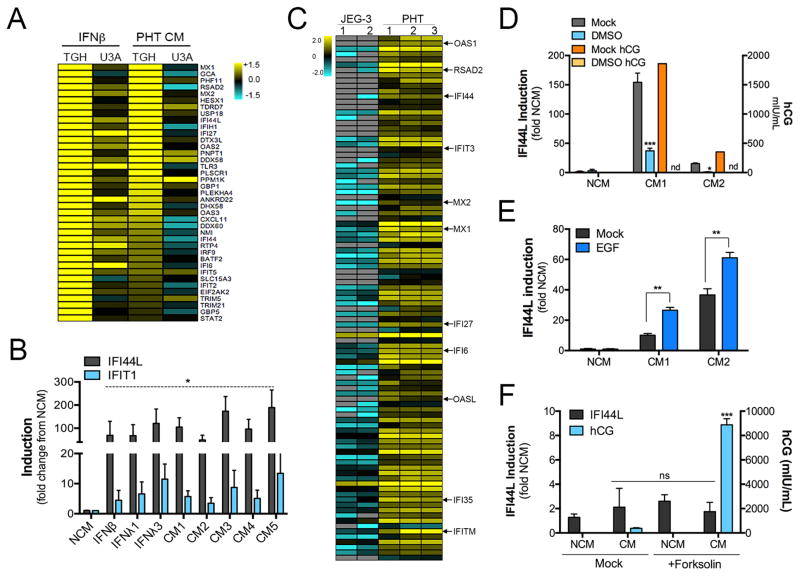
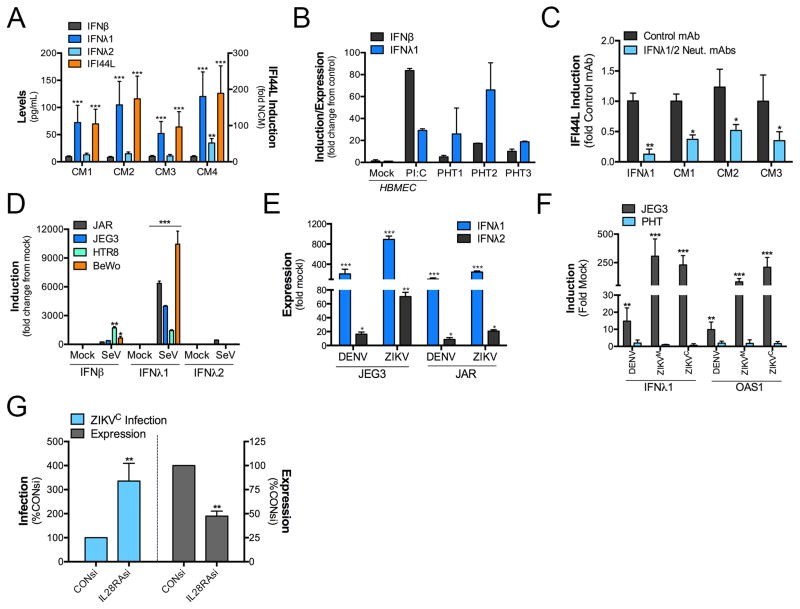
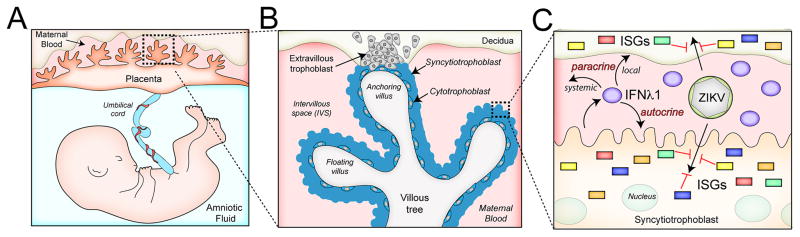
During mammalian pregnancy, the placenta acts as a barrier between the maternal and fetal compartments. The recently observed association between Zika virus (ZIKV) infection during human pregnancy and fetal microcephaly and other anomalies suggests that ZIKV may bypass the placenta to reach the fetus. This led us to investigate ZIKV infection of primary human trophoblasts (PHTs), which are the barrier cells of the placenta. We discovered that PHT cells from full-term placentas are refractory to ZIKV infection. In addition, medium from uninfected PHT cells protects non-placental cells from ZIKV infection. PHT cells constitutively release the type III interferon (IFN) IFNλ1, which functions in both a paracrine and autocrine manner to protect trophoblast and non-trophoblast cells from ZIKV infection. Our data suggest that for ZIKV to access the fetal compartment, it must evade restriction by trophoblast-derived IFNλ1 and other trophoblast-specific antiviral factors and/or use alternative strategies to cross the placental barrier.
Keywords: IFNλ; Zika virus; placenta; trophoblasts; type III interferon; virus.
Copyright © 2016 Elsevier Inc. All rights reserved.
Figures
Comment in
-
Restriction of Zika Virus by Host Innate Immunity.Cell Host Microbe. 2016 May 11;19(5):566-7. doi: 10.1016/j.chom.2016.04.019. Cell Host Microbe. 2016. PMID: 27173920
References
-
- Bazer FW, Spencer TE, Ott TL. Placental interferons. Am J Reprod Immunol. 1996;35:297–308. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
