

Activated tumor cell integrin αvβ3 cooperates with platelets to promote extravasation and metastasis from the blood stream
- PMID: 27067975
- PMCID: PMC4888909
- DOI: 10.1016/S0049-3848(16)30095-0
Activated tumor cell integrin αvβ3 cooperates with platelets to promote extravasation and metastasis from the blood stream
Abstract
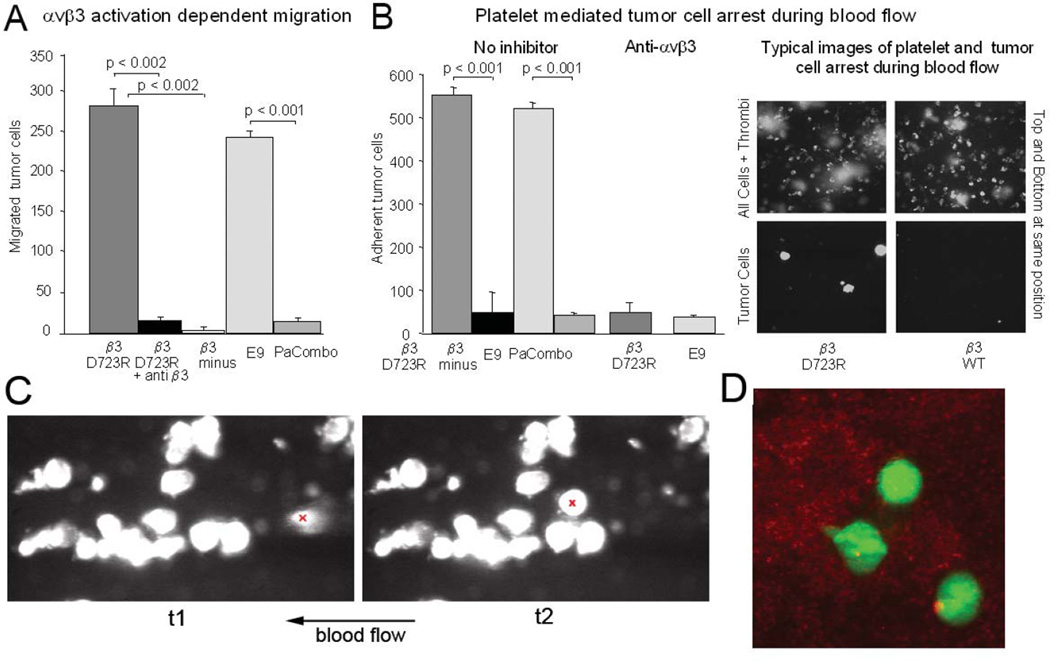
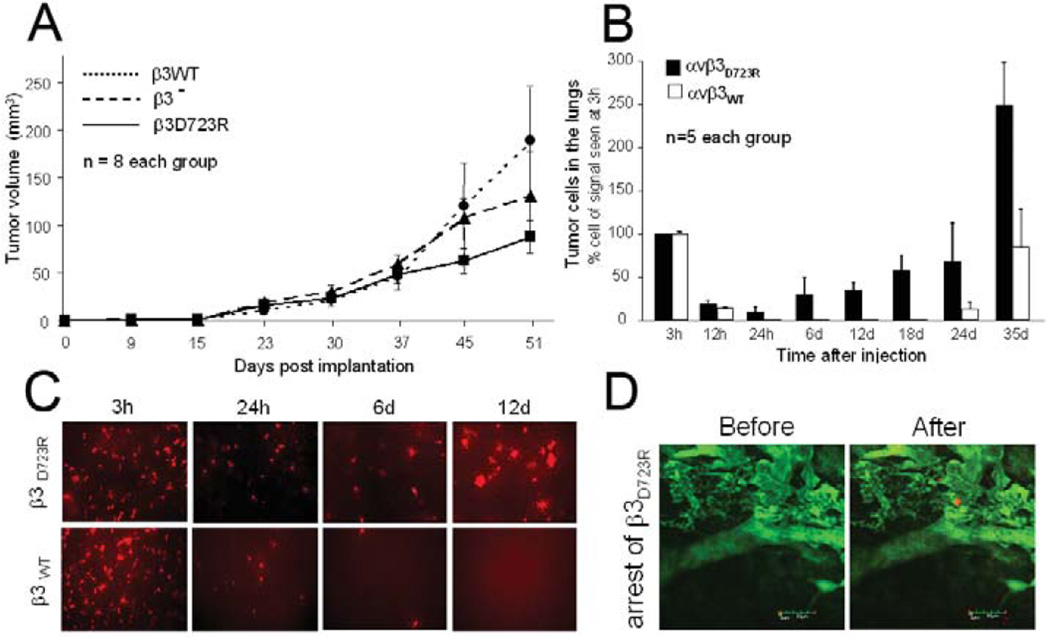
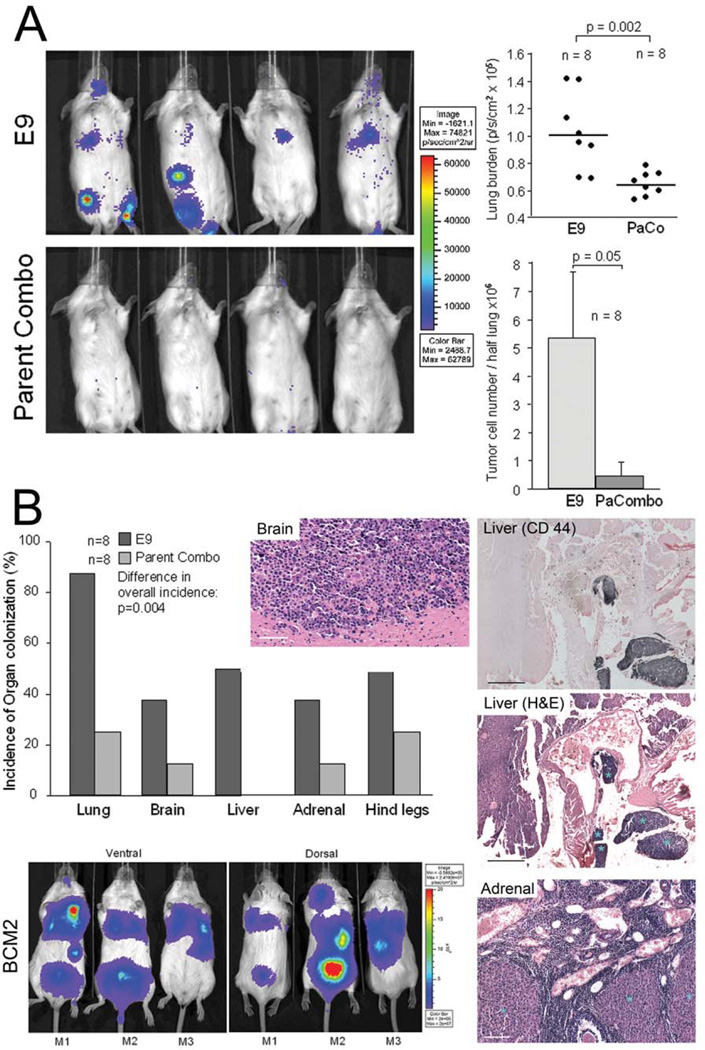
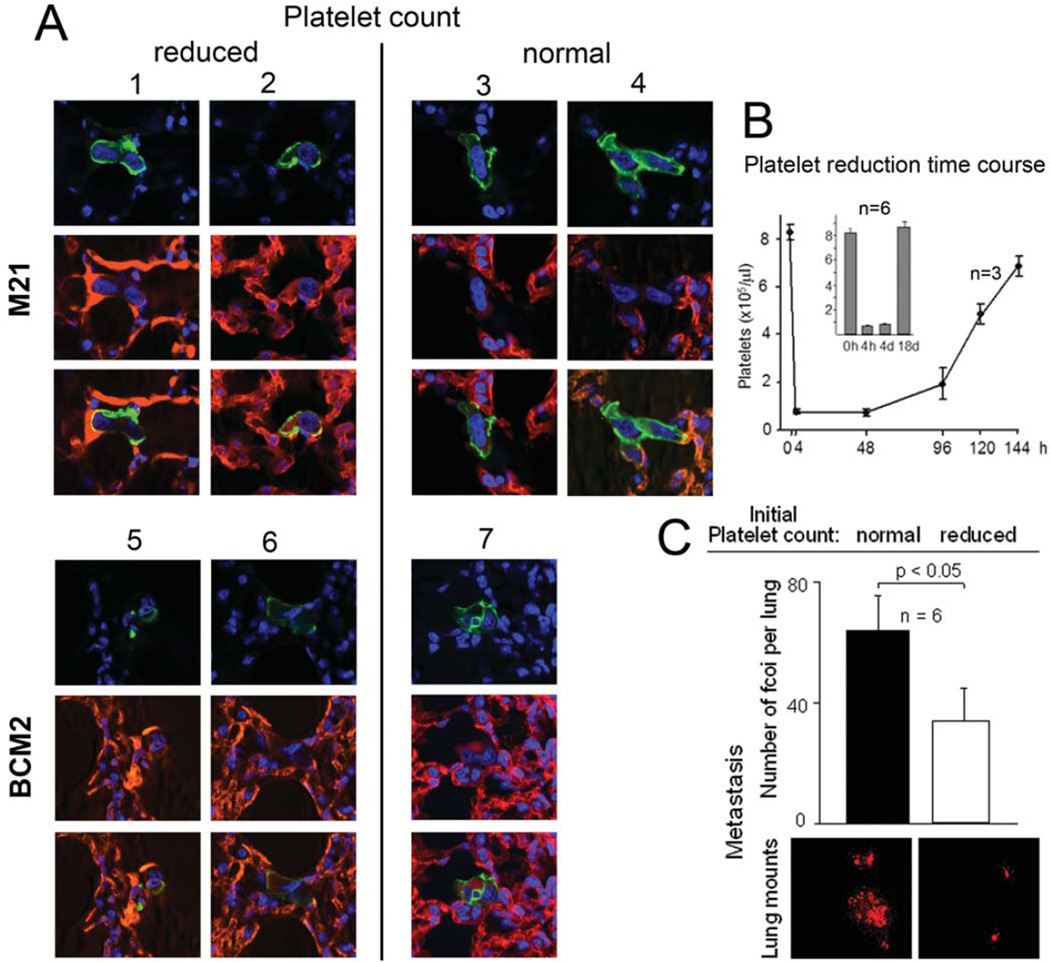
Metastasis is the main cause of death in cancer patients, and understanding mechanisms that control tumor cell dissemination may lead to improved therapy. Tumor cell adhesion receptors contribute to cancer spreading. We noted earlier that tumor cells can expressing the adhesion receptor integrin αvβ3 in distinct states of activation, and found that cells which metastasize from the blood stream express it in a constitutively high affinity form. Here, we analyzed steps of the metastatic cascade in vivo and asked, when and how the affinity state of integrin αvβ3 confers a critical advantage to cancer spreading. Following tumor cells by real time PCR, non-invasive bioluminescence imaging, intravital microscopy and histology allowed us to identify tumor cell extravasation from the blood stream as a rate-limiting step supported by high affinity αvβ3. Successful transendothelial migration depended on cooperation between tumor cells and platelets involving the high affinity tumor cell integrin and release of platelet granules. Thus, this study identifies the high affinity conformer of integrin αvβ3 and its interaction with platelets as critical for early steps during hematogenous metastasis and target for prevention of metastatic disease.
Keywords: Blood stream; Extravasation; Integrin activation; Metastasis; Platelets.
© 2016 Elsevier Ltd. All rights reserved.
Figures

Similar articles
-
Targeting activated integrin alphavbeta3 with patient-derived antibodies impacts late-stage multiorgan metastasis.Clin Exp Metastasis. 2010 Apr;27(4):217-31. doi: 10.1007/s10585-010-9320-5. Epub 2010 Mar 12. Clin Exp Metastasis. 2010. PMID: 20225083 Free PMC article.
-
Platelet ATP, Thyroid Hormone Receptor on Integrin αvβ3 and Cancer Metastasis.Horm Cancer. 2020 Feb;11(1):13-16. doi: 10.1007/s12672-019-00371-4. Epub 2019 Dec 14. Horm Cancer. 2020. PMID: 31838715 Free PMC article. Review.
-
Activated integrin alphavbeta3 cooperates with metalloproteinase MMP-9 in regulating migration of metastatic breast cancer cells.Proc Natl Acad Sci U S A. 2003 Aug 5;100(16):9482-7. doi: 10.1073/pnas.1633689100. Epub 2003 Jul 21. Proc Natl Acad Sci U S A. 2003. PMID: 12874388 Free PMC article.
-
Integrin activation controls metastasis in human breast cancer.Proc Natl Acad Sci U S A. 2001 Feb 13;98(4):1853-8. doi: 10.1073/pnas.98.4.1853. Proc Natl Acad Sci U S A. 2001. PMID: 11172040 Free PMC article.
-
Mechanistic explanation for platelet contribution to cancer metastasis.Thromb Res. 2014 May;133 Suppl 2:S149-57. doi: 10.1016/S0049-3848(14)50025-4. Thromb Res. 2014. PMID: 24862136 Review.
Cited by
-
New-generation photosensitizer-anchored gold nanorods for a single near-infrared light-triggered targeted photodynamic-photothermal therapy.Drug Deliv. 2021 Dec;28(1):1769-1784. doi: 10.1080/10717544.2021.1960923. Drug Deliv. 2021. PMID: 34470548 Free PMC article.
-
Integrin-dependent cell adhesion to neutrophil extracellular traps through engagement of fibronectin in neutrophil-like cells.PLoS One. 2017 Feb 6;12(2):e0171362. doi: 10.1371/journal.pone.0171362. eCollection 2017. PLoS One. 2017. PMID: 28166238 Free PMC article.
-
Platelet-Cancer Interplay: Molecular Mechanisms and New Therapeutic Avenues.Front Oncol. 2021 Jul 12;11:665534. doi: 10.3389/fonc.2021.665534. eCollection 2021. Front Oncol. 2021. PMID: 34322381 Free PMC article. Review.
-
High αv Integrin Level of Cancer Cells Is Associated with Development of Brain Metastasis in Athymic Rats.Anticancer Res. 2017 Aug;37(8):4029-4040. doi: 10.21873/anticanres.11788. Anticancer Res. 2017. PMID: 28739685 Free PMC article.
-
Platelets in aging and cancer-"double-edged sword".Cancer Metastasis Rev. 2020 Dec;39(4):1205-1221. doi: 10.1007/s10555-020-09926-2. Epub 2020 Sep 1. Cancer Metastasis Rev. 2020. PMID: 32869161 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
